









…やるべき事は山程ある。しかし意欲が湧いたからには仕方がない。書こう。
《前書き…というか言い訳etc》
てな訳で唐突ながら今回は、NHKスペシャル『恐竜vsほ乳類 1億5千万年の戦い』を記事にしようと思う。
先に白状しておくと、本作は筆者を恐竜の道へ引きずり込んだ元凶とも言える作品のため、折を見て記事にするつもりではあった。というのも本作はネット上において異常なほど影が薄い。『地球大進化』や『恐竜超世界』のように高く評価されることもなければ、『生命』や『恐竜絶滅 ほ乳類の戦い』のように酷評されてもいない。まるで存在自体が抹消されてしまったかのようだ。それでもネットの片隅では、本作への辛辣なツッコミ(半ばヘイトにも近い)が見受けられる。たしかにCGの出来栄えや映像の背景 ―イネ科の草原を走る恐竜たち― を指摘されては押し黙るしかない。
「だが…」
正直なところ筆者は本作『恐竜vsほ乳類』が、まずまずの良作特番だと考えている。それに言ってしまえば“恐竜vsほ乳類”は捏造でも何でもなく事実だ(#1、#α)!!…と、筆者だけが勝手に声を荒げても意味はない。そこで新シリーズでは関連した論文を織り交ぜた解説を挿みつつ『恐竜vsほ乳類』の名誉回復といこう。
《第一章・水辺の巨人〜三畳紀後期2億2000万年前》
最初の舞台となったのはアリゾナ〜ニューメキシコ州にかけて堆積したチンリ層《Chinle Formation》である。チンリ層というとアリゾナ州の「化石の森国立公園」が有名で、ここからは三畳紀後期の2億2000万年前(カーニアン〜ノーリアン)に大地を彩った多種多様な動植物の化石が発見されている(#2)。
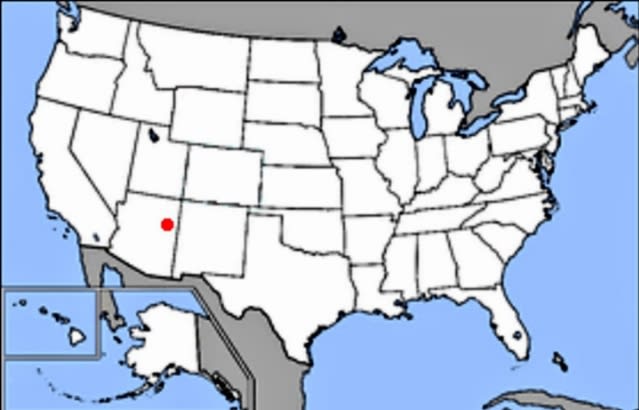
(↑Wikiコモンズより、化石の森国立公園の場所)
化石が豊かであるということは、当時の生態系を調べやすいということであり、古くからチンリ層ないし化石の森を舞台とした恐竜特番は数しれない。『恐竜vsほ乳類』もその一つだ。それでは当時のチンリ層に生息していた動物たちを軽く見ていくことにする。
作中で最初に登場したのは全長3mの水棲動物メトポサウルス《Metoposaurus》だ。メトポサウルスは由緒正しき水辺の捕食者、またの名を迷歯亜綱(めいしあこう)という巨大両生類の一派である。
古くは石炭紀から世界各地の沼や川にのさばってきた彼らは、時に地上の支配者こと単弓類とも鎬を削ってきた。そしてペルム紀末に発生した史上最悪の大量絶滅(P-T境界)をも乗り越え、乾燥化の嵐が吹き荒れる三畳紀後期にまで無事に命脈を保ってきたのである。
迷歯亜綱の特徴は貧弱な四肢 ―滅多なことには陸に上がらなかっただろう― とカスタネットのような大口だ。この口を目一杯開き、目の前を通りかかった獲物を誰彼構わず丸呑みにした(#3)。これは現代のオオサンショウウオやワニの狩りに近く、水辺を狩場とする捕食者の間では至って古典的な方法だ ―おそらく1m程度の肺魚や両生爬虫類が主食だと思われる。

(↑#Bより、メトポサウルスの生態復元)
当然メトポサウルスのような天敵がいるとなれば、近場に暮らす小動物も自衛の策を考えなければならない。

(↑Wikiコモンズより、ヴァンクレビアの産状化石)
例えば全長50cmの半水棲爬虫類であるヴァンクレビア《Vancleavea》は、全身をドラゴンさながらの鱗で覆っていた(#4)。発達した牙(本種には異歯性が見られる)から考えると、ヴァンクレビア自身も油断ならない捕食者だったのだろうが、それでも大口を開けたメトポサウルスには敵わなかったのだろう。それに一帯の捕食者は巨大な両生類だけでなく、これまた巨大な爬虫類も周囲に睨みを利かせていた。代表的なのが半水棲のフィトサウルス類で、有名どころだと全長6〜7mのルティオドン《Rutiodon》である。

(↑Wikiコモンズより、ルティオドンの組み立て骨格)
現生のワニに瓜二つなフィトサウルス類は、トングのように長い吻部と鋭い歯を用いて周囲の獲物を捕らえていた。これには明確な証拠(#5)が見つかっている。というのもマレリサウルス《Malerisaurus》と呼ばれる小型爬虫類が丸呑みにされたまま化石になっていたのだ。なお傍証にはなるが、先述のメトポサウルスとフィトサウルス類は同じ場所から発見されることが多く、互いに被食者-捕食者の関係にあったようだ(#6)。
また三畳紀の陸上には、大型動物を骨ごと噛み砕いて消化してしまう捕食者が数多くいたことが知られており(#7)、当時のオアシスは決して“水辺の楽園”というわけではなかったらしい。
そんな訳で生き残るべく恐ろしげな姿に進化したのが、映像で次に登場した全長4.5mの植物食爬虫類デスマトスクス《Desmatosuchus》である。

(↑『恐竜vsほ乳類』より、デスマトスクスの生体復元)
デスマトスクスといえば何に置いても真っ先に挙げられるのが、肩から生えた最大45cmのスパイクだ。この数本のスパイクは骨質のため非常に頑丈で、もっぱら身を守るために役立ったと考えられている。これと同様の進化を後に装盾類(アンキロサウルスやステゴサウルスの仲間)が遂げる事を考えると、両者の間には何かしらの収斂進化が働いていたのだろう。
読者の皆様がロマンたっぷりのスパイクについて知りたいのは百も承知。だがデスマトスクスの面白さはスパイクだけではない。彼らの頭部もまた、それに負けず劣らず奇妙な外形をしていた。
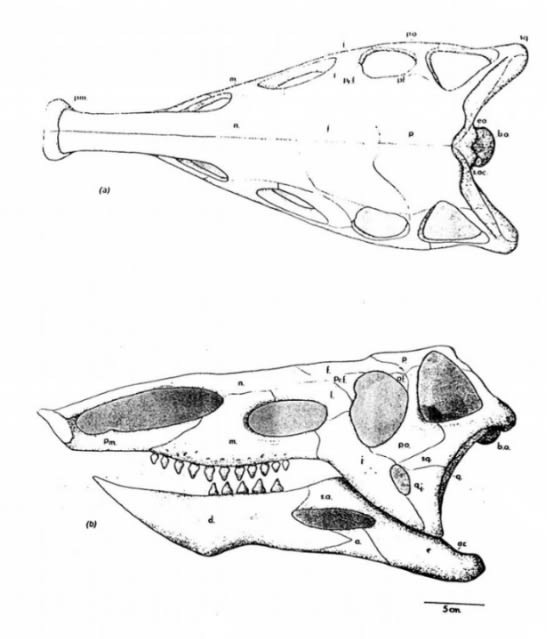
(↑#8より、図示されたデスマトスクスの頭骨。船の舳先のような形をしている)
デスマトスクスに代表されるアエトサウルス目は、揃って先端だけが潰れた口吻を備えていた。このブタのような鼻面は、さながらスコップのように使われ、地面に埋もれたトクサの地下茎などを掘り起こして食べるのに役立ったと考えられている(#8、#9)。
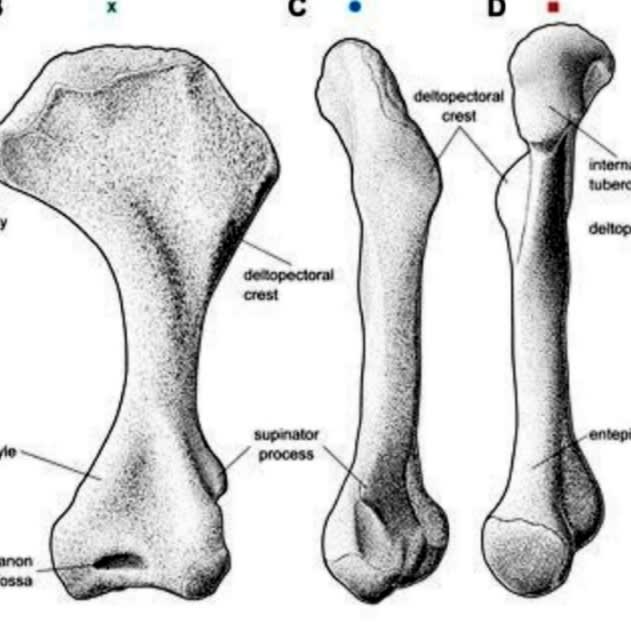
(↑#8より、アエトサウルス目の上腕骨。筋肉を付けるための凹凸が発達している)
その説を補強するように前肢の研究(#10)からも、アエトサウルス目は短く強靭な前肢で地面を掘る習性ないし能力があったことが指摘されている。もちろん作中で描写されていたように地表のシダ類をつまみ食いする事もあっただろうが、どちらかと言えば水気たっぷりの地下茎を齧るほうが好みだったのだろう。

(↑#9より、地下茎を漁るデスマトスクス)
しかしデスマトスクスには強敵が存在した。血に飢えた捕食者だけではない。真の強敵は生きたワイン樽ことプラケリアス《Placerias》だ。この全長3mの巨大生物は爬虫類ではない。無論メトポサウルスのような両生類でもなく、実は私達ほ乳類の親戚(ディキノドン類)にあたる。ディキノドン類はペルム紀後期〜三畳紀中期にかけて全盛を誇った植物食動物の仲間で、最大級のリソウィキア《Lisowicia》は体重が6トンにも及んだともされている(#11)。流石にプラケリアスの体重は1トンとされているが、それでも圧倒的な体格だ。どちらかと言えば扁平なデスマトスクスと力士よりも重厚なプラケリアス。縄張り争いでは間違いなくプラケリアスに軍配が上がったはずだ。

(↑Wikiコモンズより、プラケリアスの組み立て骨格)
さらにプラケリアスは頑強かつ強靭な頭部を備えていた。後のトリケラトプスにも似た嘴と、そこから突き出た牙状の骨(よく勘違いされているが、この部分は本物の歯ではない)。そして異様に発達した側頭窓(咬筋の通る穴)が特徴的な頭部は、見た目どおりの凄まじい力を発揮した。きっとブルドーザーよろしく土砂を掬いあげて地下茎を噛み切ったり、低木を根本から折って中の柔らかい芯を食べたりと、やりたい放題だったはずだ。
そしてもしも、プラケリアスを本気で怒らせてしまったら…。仮に飢えたフィトサウルス類が相手でも即座に踏み潰されて即刻あの世行きだろう。
ちなみにディキノドン類は鈍重そうな身体付きや発掘地の堆積物から、かつてはカバのような半水棲の動物だと考えられてきた。しかし今では骨格の再研究(#12)により、活動的な陸棲動物だったことが明らかになっている。ただしデスマトスクスのような爬虫類よりも生理機能が劣っていたため、あまり水辺を離れられなかった可能性が高い ―裏を返せば水さえ確保できれば長距離の移動も可能であり、その証拠にディキノドン類はペルム紀末〜三畳紀初頭に大繁栄していた。

(↑Wikiコモンズより、トリロフォサウルスの組み立て骨格)
大型の動物がいるなら、そこには小型の動物もいる道理。その証拠に尻尾の長いトリロフォサウルス《Trilophosaurus》や手乗りサイズのプロコロフォン科《Procolophonidae》など、実に様々な小型爬虫類がチンリ層からは見つかっている(#13)。こうした小型爬虫類たちは、前述の巨人の間を縫うようにして餌のシダやトクサを探しては、奇妙な凹凸状の歯でガジガジ食べていた ―ちょうど現生のイグアナに近いかもしれない。そんな彼らの周りには敏捷な小型の肉食爬虫類も数多く生息していた(#14)。

(↑『恐竜vsほ乳類』より、アデロバシレウスの生体復元)
最後の紹介となるのが我らが先祖のアデロバシレウス《Adelobasileus》だ。
全長は僅か10cm。作中で語られたように主に昆虫を食べていたと考えられており、現代のトガリネズミに姿も生態もそっくりだった(#15)。よく見てみると本作の哺乳類たちは、アデロバシレウスに限らず、微妙に爬行(ガニ股)しながら動き回っている。ぶっちゃけた話、ほ乳類が直立姿勢を獲得した時期は定かでない。原型は祖先とされるキノドン類など獣歯類の段階で獲得されていたものの(#16)、足全体が真下へ伸ばされた構造にはなっていなかった。それでも我々ほ乳類は中生代のどこかで爬行をやめ、真の直立歩行へ舵を切ったらしい。ただ股関節の構造などを考えるに、真の直立歩行を獲得したのは、アデロバシレウスよりずっと先 ―下手すると白亜紀までお預け― だった事は間違いない。

(↑『恐竜vsほ乳類』より、大型動物の影で生きるアデロバシレウス)
この可愛らしい動物は、化石の発掘状況がとても悲惨だったことでも知られている。なんでもアデロバシレウスの化石は、他の生物の糞化石《Coprolite》の中から見つかったらしいのだ。もし本当であれば当時の食物連鎖を伺い知る上で貴重なヒントとなるが、同時に大繁栄している今日の子孫と比べれば、本種は比較にならないほど矮小な存在と言える。とても生態系の頂点に立つ“高等動物”とは名乗れなかっただろう。
こうした様々な動物たちは、皆等しくチンリ層の厳しい環境に晒されていた。そもそも当時はパンゲア大陸全体の気温が高く、中央部は完全に砂漠化しており、緑が残るのは沿岸部など限られた地域だけだった。さらに悪いことにP-T境界による温室効果ガスの増加や、酸素濃度の急激な低下なども重なることで、我々人類からすると正気の沙汰とは思えない環境が広がる世界。それが三畳紀後期だったのだ(#17)。少なくない生物(例→テロケファルス類)が劣悪な環境に耐えられずに絶滅する中、辛うじて残っていたはそれぞれ生き残りの道を模索するしかなかった。
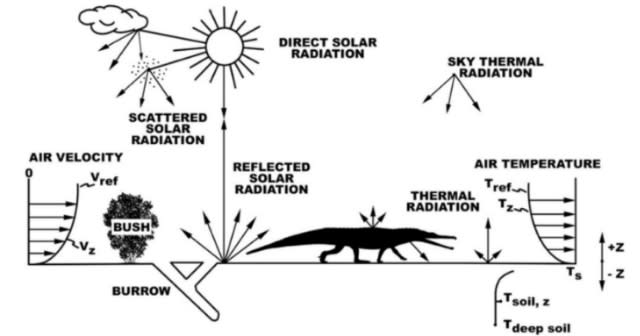
(↑#18より、当時の環境を図示したもの)
2020年に発表された熱力学的シミュレーション(#18)によると、プラケリアスやデスマトスクスのライバルこと大型の竜脚形類(例→プラテオサウルス)は、この劣悪な環境を嫌って深い森や高緯度地域へ逃れていたらしい。チンリ層は高温かつ疎林〜氾濫原および沼や川が点在する環境だったらしく、であれば竜脚形類の好みには合わなかったはずだ。またチンリ層に生息していた動物にしても、水中に潜ったり、活動時間を工夫したり、藪や巣穴に潜んだりと、何かしらの対策を身につけていたらしい。

(↑『恐竜vsほ乳類』より、干魃で息絶えたメトポサウルス(奥)と、身を潜めるアデロバシレウス)
ここまで見てきたように当時チンリ層に生息していた動物は概ねガニ股〜中腰の姿勢で動き回る長距離の移動 ―逆に咄嗟の動きは極めて敏捷だった― が苦手な種ばかりだった。これは様々な要因(進化や環境)が考えられるが、そういった生態はパンゲア超大陸の気候と相まって恐ろしい悲劇を度々もたらしたらしい。ここまでに挙げたメトポサウルス、フィトサウルス類、アエトサウルス目、ディキノドン類は、どれもボーンベッド《bone bed》が報告されている(#6、#19、#20、#21、#C)。多少の異論はあるが、おそらく動物たちは例年よりも厳しい乾季がきても馴染みの水辺を離れられず、あえなく衰弱死したものと見られている。とりわけ陸上を苦手とするメトポサウルスにとっては命取りとなったことだろう。同様の現象は太古の昔に限った話ではなく、現代でも頻繁に報告されている(#β、#γ)。
こうした生物や環境に囲まれながら、ほ乳類アデロバシレウスは産声をあげた。
しかし今でこそ世界を統べる王の器と讃えられる哺乳類でさえ、当時はまだ新参者のぺーペーでしかない事を、読者の皆様は次回以降に知ることになる。
《参考文献》
[論文]
#1『Dinosaur physiology and the origin of mammals』(恐竜vsほ乳類)
#2『Vertebrate trace fossils from New Mexico and their significance'』(ニューメキシコの古脊椎動物)
#3『How tetrapods feed in water: a functional analysis by paradigm』(半水棲捕食者の戦術)
#4『The osteology and relationships of Vancleavea campi (Reptilia: Archosauriformes)』(ヴァンクレビアの骨学)
#5『Malerisaurus, a new eosuchian reptile from the Late Triassic of India』(フィトサウルス類の餌食)
#6『Mortality dynamics and fossilisation pathways of a new metoposaurid-dominated multitaxic bonebed from India: a window into the Late Triassic vertebrate palaeoecosystem』(両生爬虫類の集団化石)
#7『Coprolites of Late Triassic carnivorous vertebrates from Poland: an integrative approach』(ポーランド産の糞化石)
#8『The Triassic thecodontian reptile Desmatosuchus: osteology and relationships』(デスマトスクスの骨学)
#9『Aetosauria: a clade of armoured pseudosuchians from the Late Triassic continental beds』(アエトサウルス類の食性と分類)
#10『Osteology of a forelimb of an aetosaur Stagonolepis olenkae (Archosauria: Pseudosuchia: Aetosauria) from the Krasiejów locality in Poland and its probable adaptations for a …』(アエトサウルス類の前肢)
#11『Resizing Lisowicia bojani: volumetric body mass estimate and 3D reconstruction of the giant Late Triassic dicynodont』(リソウィキアの体重)
#12『The aquatic Lystrosaurus: A palaeontological myth』(ディキノドン類は半水棲だった?)
#13『A new procolophonid and a new tetrapod of uncertain, possibly procolophonian affinities from the Upper Triassic of Virginia』(三畳紀の小型爬虫類)
#14『The origin of the crocodiloid tarsi and the interrelationships of thecodontian archosaurs.』(小型の肉食爬虫類)
#15『Adelobasileus from the Upper Triassic of West Texas: the oldest mammal』(アデロバシレウスの論文)
#16『Reassessment of the postcranial anatomy of Prozostrodon brasiliensis and implications for postural evolution of non-mammaliaform cynodonts』(直立姿勢の獲得)
#17『The latitudinal diversity gradient of tetrapods across the Permo-Triassic mass extinction and recovery interval』(超大陸パンゲアの気候)
#18『Modeling Dragons: Using linked mechanistic physiological and microclimate models to explore environmental, physiological, and morphological constraints on the early evolution of dinosaurs』(獣脚類と竜脚形類の代謝機能)
#19『Taphonomy of the Lamy amphibian quarry: a Late Triassic bonebed in New Mexico, USA』(メトポサウルスの墓場?)
#20『Taphonomy and Depositional Setting of the Placerias Quarry (Chinle Formation: Late Triassic, Arizona)』(旱魃の犠牲となったプラケリアス)
#21『New specimen of Dinodontosaurus (Therapsida, Anomodontia) from west-central Argentina (Chañares Formation) and a reassessment of the Triassic Dinodontosaurus Assemblage Zone of …』(ディノドントサウルスの集団化石)
[洋書]
#A『The Triassic-Jurassic Terrestrial Transition: 37』(三畳紀〜ジュラ紀の諸々)
#B『Petrified Forest National Park: A Wilderness Bound in Time』(化石の森国立公園)
#C『Taphonomy of the Late Triassic Placerias Quarry (Petrified Forest Member, Chinle Formation) of eastern Arizona』(化石の森におけるタフォノミー)
[ネット記事]
#α『How the earliest mammals thrived alongside dinosaurs』(恐竜と哺乳類は共存した)(Nature)
#β『野生の馬が大量死、熱波で水場干上がる オーストラリア』(CNNニュース)
#γ『池でフナやコイが大量死 猛暑が原因か 千葉』(0テレNews)
[書籍]
・恐竜vsほ乳類(書籍版)
・恐竜学入門-かたち・生態・絶滅
・恐竜異説
・ホルツ博士の最新恐竜事典
・愛しのブロントサウルス
・オーロラをみた恐竜たち(図録)
・地球最古の恐竜展(図録)
《関連記事》
《ネタ元》










