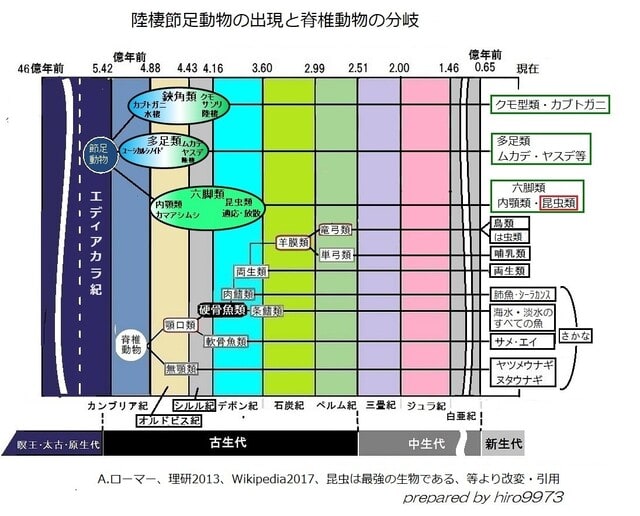
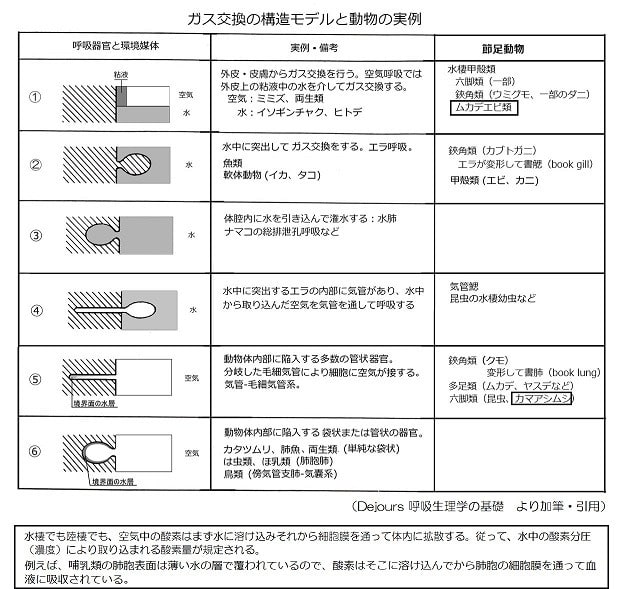
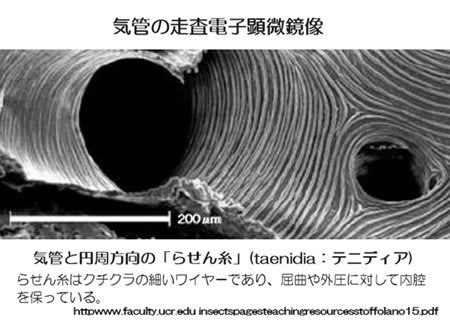
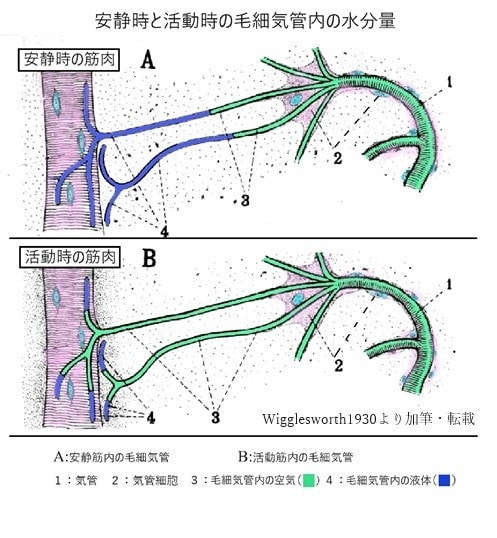
両生類の呼吸―その2
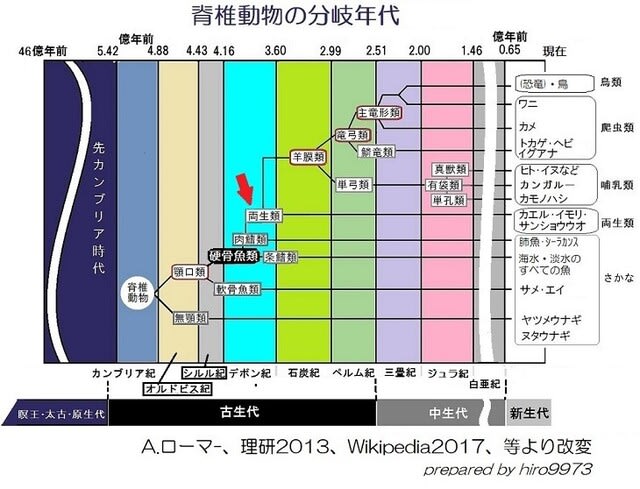
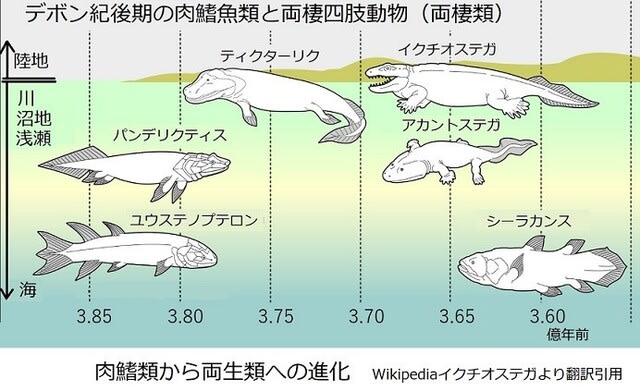
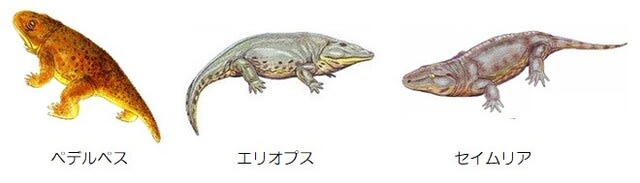
前回の記事のようにデボン紀後期に肉鰭類は陸上進出していきました。そのような進化への道を進んだ要因について考えてみました。
海の中は、魚たちにとって温度変化が小さくて、エラから十分に酸素を吸収できて、食べ物も十分にある上に、浮力で体が支えられているような快適な環境でした。
一方進出を試みた陸上は、肺や皮膚で呼吸ができたとしても、変温動物なので低い気温の時には動きにくくなり(攻撃を受けやすい)、体表と肺を使う呼吸のために体の水分が蒸発して干からびてしまう恐れがあり、さらに浮力がなくなるので移動のためには水中よりも多くのエネルギーが必要、など多く困難がありました。
ローマーは「脊椎動物の歴史」の中で、「両生類が出現したデボン紀は季節的な干ばつが繰り返された時代であった。しばしば川のながれはとまり、残された水たまりには魚類や初期の両生類たちが殺到し、よどんでいた。・・・肺を持っていたので空気を呼吸できた。・・・陸上歩行に適した四肢を持っていたので川や池の水たまりにたどり着き・・・」と
厳しい環境による進化圧力により水陸両用となったとしています。
ローマー以降、魚の体の構造を陸上生活に適応するように作り変えるという個々の構造の進化については多くの遺伝子研究などで解明が進んでいます。
また、最近までに環境や酸素濃度について多くの知見が積み上げられました。
そこで、2千万年以上にわたり魚たちを陸上への進出に駆り立てた状況や動機について、陸上への進出が始まった3億7千万年~3億6千万年前の時代を次の3つの視点で調べてみました。
A)環境の激変
B)酸素濃度
C)エラから肺への飛躍
A)環境の激変
A-1)大規模火山噴火
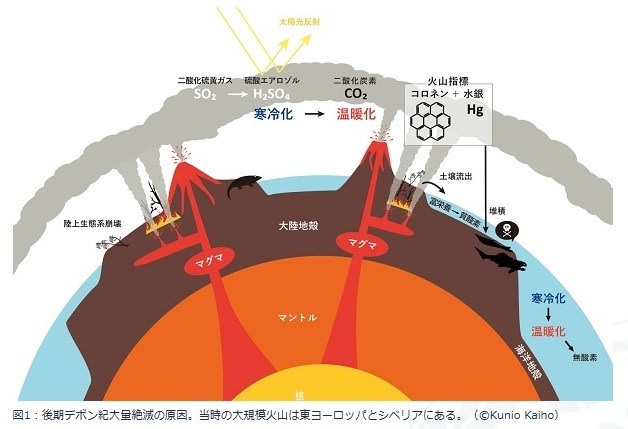
3億7千万年前のデボン紀後期に、全海洋生物種のうち82 %が絶滅した生物大量絶滅がおきました。地質の堆積記録では複数の大きな環境変動が示されています。
2021年に東北大学理学部からこの大量絶滅の原因は、大規模火山噴火であるとする報告が発表されました。
そのシナリオ:
① 地球の深部でおきる大規模なマグマの活動(スーパープルーム)は地殻変動や大規模火山噴火をおこします。そのときに地殻内の石油や石炭の有機物が加熱されて特有の有機分子(コロネン)や、二酸化硫黄(SO2)、二酸化炭素(CO2)、メタンが噴出します。
② 二酸化硫黄は大気中で硫酸エアロゾル(空気中に硫酸の微粒子が多数浮かんだ状態)を作って雲のように太陽光を遮断するので地表は寒冷化します。その後エアロゾルは雨や拡散により大気中から失われると、それまで寒冷化していた地表は二酸化炭素とメタンによる温室効果で急速に温暖化することになります。
③ 温暖化によって、深海に酸素を運ぶ深層海流が弱くなります。
まず、深層海流についてですが、大西洋のメキシコ湾から北上してグリーンランド沖に流れ込むメキシコ湾流は、そこで冷やされて密度が高くなり、さらに塩分濃度も上がります。重くなった海水は深海へ沈み込む海流となって大西洋の深海を南に進みインド洋から太平洋へと熱や物質、特に酸素を行き渡らせるという深海の大循環のことです。
さて、極端な温暖化が生じると海水温の上昇で密度が小さくなり、それに加えて海氷から溶けた真水が混ざってさらに密度が小さくなります。そのため軽くなった海水はグリーンランド沖の深海への沈み込みが起きなくなって世界中の深海に酸素が行き渡らない無酸素状態となります。
こうして、大規模なマグマの活動がもたらす急速な寒冷化とその後の温暖化という大規模な気候変動は、大気の気温を上昇させて中~深海の無酸素状態を生じさせるために深海の生物の大量絶滅が起こったと考えられています。(Global and Planetary Change 2021/2/20)
この事象は、数万年から数百万年かけて徐々に回復していくようです。
A-2)植物の進化
植物はシルル紀後期からの進化によってデボン紀後期には高さ30mを超えるシダ類のリンボク(ヒカゲノカズラ類)や20m以上のロボク(トクサ類)などの胞子植物、さらに種子で増える種子植物も出現してこれらによる最初の森が出現しました。
 藻類や樹木がそれまで一切の生物のいなかった荒地に徐々に拡大していくに従って、雨水により植物や藻類の窒素化合物やリンなどの栄養分と土壌のミネラルは河川を通って海に流入しました。
藻類や樹木がそれまで一切の生物のいなかった荒地に徐々に拡大していくに従って、雨水により植物や藻類の窒素化合物やリンなどの栄養分と土壌のミネラルは河川を通って海に流入しました。
シアノバクテリアや水中のプランクトンなどは、この大量の栄養塩と海水中に溶けている酸素を使って急速に繁殖(赤潮の発生)すると海水中の酸素量は著しく低下し、さらにバクテリアによるその死骸の分解でも酸素を消費するためにいっそう低酸素となって、浅い海水中では酸素の乏しい状態(貧酸素状態)が生じたとされています。(日本地質学会HP 海洋無酸素事変より)
このように、大規模火山噴火と植物の進出という環境の変化により浅い海から深海にまで及ぶ貧酸素~無酸素の状態が引き起こされたために前海洋生物種の80%以上が絶滅したと考えられています。
B)空気中の酸素濃度の変化
デボン紀初期の4億1千万年前に約25%あった空気中の酸素濃度は3億9千万年前のデボン紀中期には約13%まで低下したと推測されています。
デボン紀後期に植物の大躍進が始まると大気中の二酸化炭素が吸収されて、その炭素から樹木が作られると同時に、排出された酸素は大気に蓄積されていきました。こうして、デボン紀後期から森林時代の温暖湿潤な石炭紀を経て、ペルム紀初期(2億9千万年前)に至る1億年間に空気中の酸素濃度は約30~35%まで上昇していきました。現在の21%よりもはるかに高濃度になっていったのです。魚たちが陸上に進出を始めた頃には17~18%程度まで上がっていました。

一方、二酸化炭素はデボン紀中頃に現在の15倍(0.04%X15=0.6%)もあって温室効果により温暖(むしろ暑い?)な環境を作っていましたが、大森林の樹木が倒れ埋没すると石炭となって大気から取り除かれたので、石炭紀初期には現在と同じ0.04%程度まで低下し気候は次第に寒冷化していきました。
C)エラから肺への切り替えという飛躍
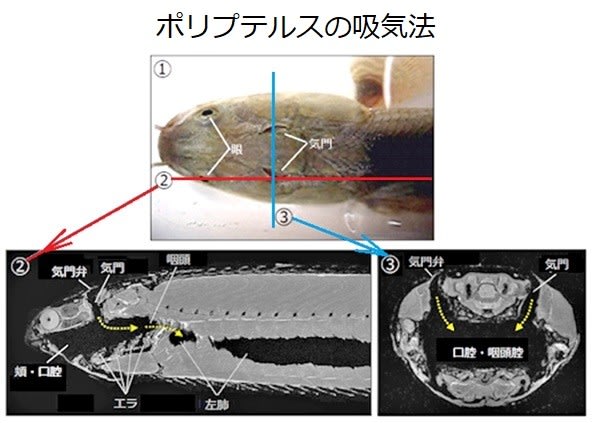
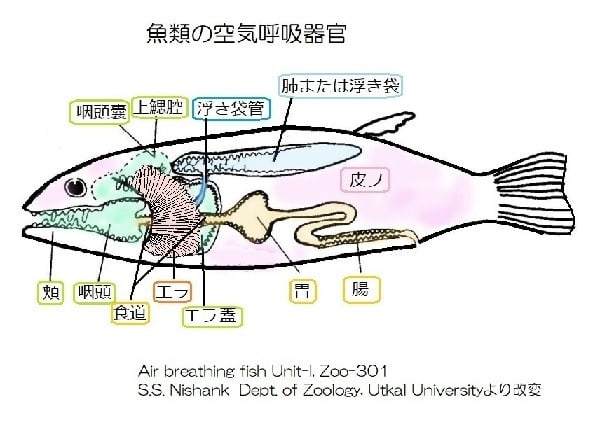
現生の魚たち(条鰭類)の中には、肺を使って陸上生活の可能なポリプテルスがいます。
肺はすでにシルル紀中期に初期の条鰭類が獲得していました。スズキ目の魚では口腔や咽頭とエラ蓋に補助呼吸器を発達させています。
・スズキ目のトビハゼの仲間(ムツゴロウなど)は、表皮に乾燥を防ぐ薄い水の膜があって、必要な酸素の約80%は表皮を透過して皮下の毛細血管で吸収しています。さらに口腔、咽頭、えら蓋の内側の空間に水と空気で泡を作り、そこから内腔表面に張り巡らされた毛細血管網の血液に酸素が吸収されています。
・スズキ目のキノボリウオの仲間:アナバス、ライギョ、ベタなど
実際は木に登ることはなく、雨天時などに地面を這い回る程度です。えらの上の頭蓋内に上鰓器官(じょうさいきかん:ラビリンス器官)という迷宮のような構造を作って内部の表面積を拡大し、その表面の網状の毛細血管から空気呼吸をします。補助的な呼吸器官ですが、陸上でも湿った環境なら長時間生存できます。
・ポリプテルス:肉鰭類に似たぶ厚いヒレを持っています。
マギール大学(カナダ)では、ほぼ1年間陸上でポリプテルスを飼育し交配させたところ、幼魚が成長する間に胸の骨格とそれを支える支持が強くなり、歩行中に頭蓋骨を持ち上げて頭/首の動きが大きくなるという変化が起きました(発育における可塑性)。
 その結果、胸ヒレを体に近づけ、ヒレが滑らずに歩くようになりました。これは魚が最初にヒレを持って陸地を歩いたときに起こった変化を反映していると推測されています。
その結果、胸ヒレを体に近づけ、ヒレが滑らずに歩くようになりました。これは魚が最初にヒレを持って陸地を歩いたときに起こった変化を反映していると推測されています。
このように、現生の魚が示す空気呼吸への適応をみてみると、エラ呼吸から肺呼吸への進化は特別に困難な飛躍ではなかったようです。
(もちろん陸上での活発な生活のためには四肢に加えて、緻密心筋や心臓にもっと酸素を送れるような循環器の構造的な進化が必要です)
(詳しくは水中の動物たちの呼吸15,16,17を参照してください)
○まとめ:魚たちが飽くことなく陸上への進出に挑んだ理由について
デボン紀後半の時期に大規模火山活動と植物の陸上進出とが起きたために海の中の酸素が著しく減少する海洋無酸素事変が生じて、魚たちが暮らす海中の酸素が減っていきました。
一方、陸上では空中の酸素濃度が上昇の一途をたどっている上に、気候は二酸化炭素による温室効果のために温暖で高湿度でした。変温動物の魚たちにとって、陸上は空気呼吸という問題を解決できればとても魅力的な新天地になっていたのです。
さて、魚たちにとってエラ呼吸から肺呼吸への切り替えは決して大きな障壁でなかったようです。現生のポリプテルスは肺を使い、スズキ目では頭蓋内の補助呼吸器を使って、どちらも魚の体制のまま陸上を這いまわっています。空気呼吸は四肢や胸郭を陸上用に改変するよりも容易に獲得できる機能と思われました。
このように海中と陸上の環境が変化して空気呼吸への進化を促す条件が揃うと、魚たちは肺と皮膚を使って(果敢に!)陸上へと進出していったのでしょう。
両生類への進化がはじまったのです。
ところで、多くの成書や論文報告では水中でヒレが脚に変わってから陸に上がったとされていますが、これまでの概観から私にはポリプテルスのように、まず魚の体制のまま肺呼吸をして陸に上がり、それから脚などが進化していったのではないかと思われました。
参考文献
A.ローマー. 脊椎動物の歴史 どうぶつ社 東京1991
PD.ウォード.恐竜はなぜ鳥に進化したか 文藝春秋 東京 2008
D.プロセロ.化石が語る生命の歴史 11の化石・生命誕生を語る・古生代 築地書館
東京 2018
東北大学理学部プレスリリース デボン紀の大量絶滅は大規模火山活動が原因
2021年2月22日 |
日本地質学会HP 海洋無酸素事変
大河内直彦 海洋無酸素事変 科学 vol80 No11 2010
Wikipedia 植物の進化より図の引用
EM. Standen. Developmental plasticity and the origin of tetrapods
Nature;513, 54–58 (2014)
ポリプテルスの歩行図は以下より転載https://www.youtube.com/watch?v=mKxRe0hAQmg&t=8s
前回の記事のようにデボン紀後期に肉鰭類は陸上進出していきました。そのような進化への道を進んだ要因について考えてみました。
海の中は、魚たちにとって温度変化が小さくて、エラから十分に酸素を吸収できて、食べ物も十分にある上に、浮力で体が支えられているような快適な環境でした。
一方進出を試みた陸上は、肺や皮膚で呼吸ができたとしても、変温動物なので低い気温の時には動きにくくなり(攻撃を受けやすい)、体表と肺を使う呼吸のために体の水分が蒸発して干からびてしまう恐れがあり、さらに浮力がなくなるので移動のためには水中よりも多くのエネルギーが必要、など多く困難がありました。
ローマーは「脊椎動物の歴史」の中で、「両生類が出現したデボン紀は季節的な干ばつが繰り返された時代であった。しばしば川のながれはとまり、残された水たまりには魚類や初期の両生類たちが殺到し、よどんでいた。・・・肺を持っていたので空気を呼吸できた。・・・陸上歩行に適した四肢を持っていたので川や池の水たまりにたどり着き・・・」と
厳しい環境による進化圧力により水陸両用となったとしています。
ローマー以降、魚の体の構造を陸上生活に適応するように作り変えるという個々の構造の進化については多くの遺伝子研究などで解明が進んでいます。
また、最近までに環境や酸素濃度について多くの知見が積み上げられました。
そこで、2千万年以上にわたり魚たちを陸上への進出に駆り立てた状況や動機について、陸上への進出が始まった3億7千万年~3億6千万年前の時代を次の3つの視点で調べてみました。
A)環境の激変
B)酸素濃度
C)エラから肺への飛躍
A)環境の激変
A-1)大規模火山噴火
3億7千万年前のデボン紀後期に、全海洋生物種のうち82 %が絶滅した生物大量絶滅がおきました。地質の堆積記録では複数の大きな環境変動が示されています。
2021年に東北大学理学部からこの大量絶滅の原因は、大規模火山噴火であるとする報告が発表されました。
そのシナリオ:
① 地球の深部でおきる大規模なマグマの活動(スーパープルーム)は地殻変動や大規模火山噴火をおこします。そのときに地殻内の石油や石炭の有機物が加熱されて特有の有機分子(コロネン)や、二酸化硫黄(SO2)、二酸化炭素(CO2)、メタンが噴出します。
② 二酸化硫黄は大気中で硫酸エアロゾル(空気中に硫酸の微粒子が多数浮かんだ状態)を作って雲のように太陽光を遮断するので地表は寒冷化します。その後エアロゾルは雨や拡散により大気中から失われると、それまで寒冷化していた地表は二酸化炭素とメタンによる温室効果で急速に温暖化することになります。
③ 温暖化によって、深海に酸素を運ぶ深層海流が弱くなります。
まず、深層海流についてですが、大西洋のメキシコ湾から北上してグリーンランド沖に流れ込むメキシコ湾流は、そこで冷やされて密度が高くなり、さらに塩分濃度も上がります。重くなった海水は深海へ沈み込む海流となって大西洋の深海を南に進みインド洋から太平洋へと熱や物質、特に酸素を行き渡らせるという深海の大循環のことです。
さて、極端な温暖化が生じると海水温の上昇で密度が小さくなり、それに加えて海氷から溶けた真水が混ざってさらに密度が小さくなります。そのため軽くなった海水はグリーンランド沖の深海への沈み込みが起きなくなって世界中の深海に酸素が行き渡らない無酸素状態となります。
こうして、大規模なマグマの活動がもたらす急速な寒冷化とその後の温暖化という大規模な気候変動は、大気の気温を上昇させて中~深海の無酸素状態を生じさせるために深海の生物の大量絶滅が起こったと考えられています。(Global and Planetary Change 2021/2/20)
この事象は、数万年から数百万年かけて徐々に回復していくようです。
A-2)植物の進化
植物はシルル紀後期からの進化によってデボン紀後期には高さ30mを超えるシダ類のリンボク(ヒカゲノカズラ類)や20m以上のロボク(トクサ類)などの胞子植物、さらに種子で増える種子植物も出現してこれらによる最初の森が出現しました。
藻類や樹木がそれまで一切の生物のいなかった荒地に徐々に拡大していくに従って、雨水により植物や藻類の窒素化合物やリンなどの栄養分と土壌のミネラルは河川を通って海に流入しました。シアノバクテリアや水中のプランクトンなどは、この大量の栄養塩と海水中に溶けている酸素を使って急速に繁殖(赤潮の発生)すると海水中の酸素量は著しく低下し、さらにバクテリアによるその死骸の分解でも酸素を消費するためにいっそう低酸素となって、浅い海水中では酸素の乏しい状態(貧酸素状態)が生じたとされています。(日本地質学会HP 海洋無酸素事変より)
このように、大規模火山噴火と植物の進出という環境の変化により浅い海から深海にまで及ぶ貧酸素~無酸素の状態が引き起こされたために前海洋生物種の80%以上が絶滅したと考えられています。
B)空気中の酸素濃度の変化
デボン紀初期の4億1千万年前に約25%あった空気中の酸素濃度は3億9千万年前のデボン紀中期には約13%まで低下したと推測されています。
デボン紀後期に植物の大躍進が始まると大気中の二酸化炭素が吸収されて、その炭素から樹木が作られると同時に、排出された酸素は大気に蓄積されていきました。こうして、デボン紀後期から森林時代の温暖湿潤な石炭紀を経て、ペルム紀初期(2億9千万年前)に至る1億年間に空気中の酸素濃度は約30~35%まで上昇していきました。現在の21%よりもはるかに高濃度になっていったのです。魚たちが陸上に進出を始めた頃には17~18%程度まで上がっていました。
一方、二酸化炭素はデボン紀中頃に現在の15倍(0.04%X15=0.6%)もあって温室効果により温暖(むしろ暑い?)な環境を作っていましたが、大森林の樹木が倒れ埋没すると石炭となって大気から取り除かれたので、石炭紀初期には現在と同じ0.04%程度まで低下し気候は次第に寒冷化していきました。
C)エラから肺への切り替えという飛躍
現生の魚たち(条鰭類)の中には、肺を使って陸上生活の可能なポリプテルスがいます。
肺はすでにシルル紀中期に初期の条鰭類が獲得していました。スズキ目の魚では口腔や咽頭とエラ蓋に補助呼吸器を発達させています。
・スズキ目のトビハゼの仲間(ムツゴロウなど)は、表皮に乾燥を防ぐ薄い水の膜があって、必要な酸素の約80%は表皮を透過して皮下の毛細血管で吸収しています。さらに口腔、咽頭、えら蓋の内側の空間に水と空気で泡を作り、そこから内腔表面に張り巡らされた毛細血管網の血液に酸素が吸収されています。
・スズキ目のキノボリウオの仲間:アナバス、ライギョ、ベタなど
実際は木に登ることはなく、雨天時などに地面を這い回る程度です。えらの上の頭蓋内に上鰓器官(じょうさいきかん:ラビリンス器官)という迷宮のような構造を作って内部の表面積を拡大し、その表面の網状の毛細血管から空気呼吸をします。補助的な呼吸器官ですが、陸上でも湿った環境なら長時間生存できます。
・ポリプテルス:肉鰭類に似たぶ厚いヒレを持っています。
マギール大学(カナダ)では、ほぼ1年間陸上でポリプテルスを飼育し交配させたところ、幼魚が成長する間に胸の骨格とそれを支える支持が強くなり、歩行中に頭蓋骨を持ち上げて頭/首の動きが大きくなるという変化が起きました(発育における可塑性)。
その結果、胸ヒレを体に近づけ、ヒレが滑らずに歩くようになりました。これは魚が最初にヒレを持って陸地を歩いたときに起こった変化を反映していると推測されています。 このように、現生の魚が示す空気呼吸への適応をみてみると、エラ呼吸から肺呼吸への進化は特別に困難な飛躍ではなかったようです。
(もちろん陸上での活発な生活のためには四肢に加えて、緻密心筋や心臓にもっと酸素を送れるような循環器の構造的な進化が必要です)
(詳しくは水中の動物たちの呼吸15,16,17を参照してください)
○まとめ:魚たちが飽くことなく陸上への進出に挑んだ理由について
デボン紀後半の時期に大規模火山活動と植物の陸上進出とが起きたために海の中の酸素が著しく減少する海洋無酸素事変が生じて、魚たちが暮らす海中の酸素が減っていきました。
一方、陸上では空中の酸素濃度が上昇の一途をたどっている上に、気候は二酸化炭素による温室効果のために温暖で高湿度でした。変温動物の魚たちにとって、陸上は空気呼吸という問題を解決できればとても魅力的な新天地になっていたのです。
さて、魚たちにとってエラ呼吸から肺呼吸への切り替えは決して大きな障壁でなかったようです。現生のポリプテルスは肺を使い、スズキ目では頭蓋内の補助呼吸器を使って、どちらも魚の体制のまま陸上を這いまわっています。空気呼吸は四肢や胸郭を陸上用に改変するよりも容易に獲得できる機能と思われました。
このように海中と陸上の環境が変化して空気呼吸への進化を促す条件が揃うと、魚たちは肺と皮膚を使って(果敢に!)陸上へと進出していったのでしょう。
両生類への進化がはじまったのです。
ところで、多くの成書や論文報告では水中でヒレが脚に変わってから陸に上がったとされていますが、これまでの概観から私にはポリプテルスのように、まず魚の体制のまま肺呼吸をして陸に上がり、それから脚などが進化していったのではないかと思われました。
参考文献
A.ローマー. 脊椎動物の歴史 どうぶつ社 東京1991
PD.ウォード.恐竜はなぜ鳥に進化したか 文藝春秋 東京 2008
D.プロセロ.化石が語る生命の歴史 11の化石・生命誕生を語る・古生代 築地書館
東京 2018
東北大学理学部プレスリリース デボン紀の大量絶滅は大規模火山活動が原因
2021年2月22日 |
日本地質学会HP 海洋無酸素事変
大河内直彦 海洋無酸素事変 科学 vol80 No11 2010
Wikipedia 植物の進化より図の引用
EM. Standen. Developmental plasticity and the origin of tetrapods
Nature;513, 54–58 (2014)
ポリプテルスの歩行図は以下より転載https://www.youtube.com/watch?v=mKxRe0hAQmg&t=8s