








昆虫の呼吸-11
前回は細胞に直接空気を供給する気管―毛細気管呼吸を行う昆虫には巨大化に制限がかかるという話でした。細胞数が相似比の3乗に比例して増えること(例えば体長が10倍になれば細胞数は1000倍になること)により気管・毛細気管という空気配管の体積が昆虫体内に占める割合が著しく大きくなることがその要因と考えました。一方、私たち脊椎動物はガス交換のために循環系という血液配管を持つことで巨大化に伴う細胞への酸素供給の問題を解決しています。
今回は気管-毛細気管の構造と水分の移動についてです。
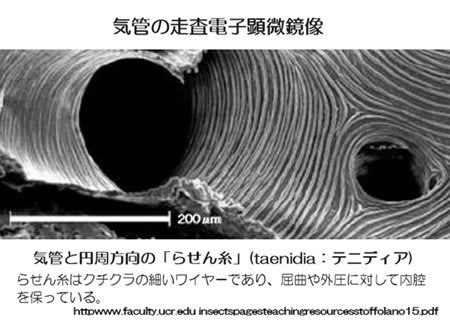
気管は筋肉の収縮や体内の圧力の上昇で圧縮・虚脱し、圧力が弱まるとらせん糸(テニディア)の弾性で拡張するという機械的な運動で換気しています(次図)。一方、毛細気管にはテニディアはなくて直径は1~0.2μmと細く呼吸のためのガス交換は拡散で行われています。(昆虫の呼吸-5、6参照)。その壁の厚さはわずかに0.01~0.03μm(Weis-Foghの原文では100 ~ 300 オングストローム)であり酸素や二酸化炭素の透過にはほとんど影響しないと考えられています。ちなみに酸素分子の大きさは3~4オングストロームです。
この図では直径100μmと150μmの気管と密集して気管を内張しているテニディアが見られます。
毛細気管内の水分移動と酸素拡散
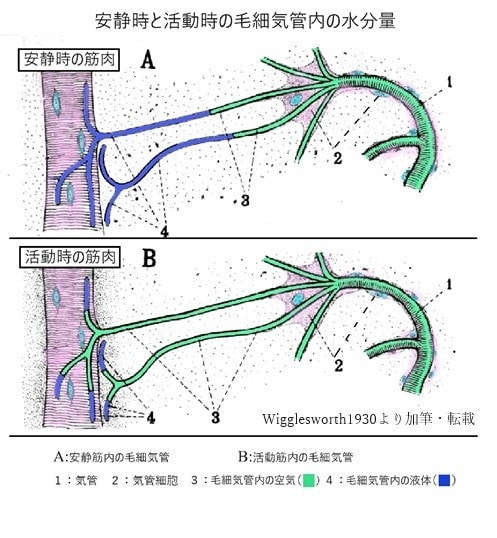
1930年にWigglesworthは蚊の幼虫の観察・実験を基に、毛細気管が細胞内に陥入している抹消では毛細気管内は細胞から浸出した液体で満たされていることを指摘しています。彼の実験では以下のことがわかっています。
・安静時には末梢部分は液体で満たされている(図A)
・窒息や運動中の組織液中には過剰な乳酸がある
・窒息あるいは活動している筋肉では、毛細気管中の液体は吸収されて毛細気管内
の空気が細胞に向かって伸びていく(図B)
・幼虫に塩化ナトリウムと乳酸カリウムを与えて高浸透圧にすると筋肉の活動時と同じ様な液体の吸収と空気柱の伸張がみられる。
これらの結果から、
毛細気管が陥入している細胞質と周囲の組織液は筋肉細胞の活動のために、乳酸やその他の濃度が高くなって、浸透圧が上昇する。
その結果、毛細気管のなかの浸出液は組織中に再吸収されて空気が毛細気管の末梢まで届くようになる。
このように、浸透圧作用により活動時や低酸素時には毛細気管の末梢にまで空気が入って効率よく細胞に酸素が行き渡ると考察しています。
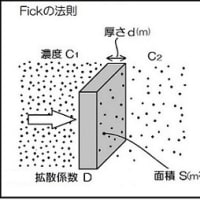
次の表に、酸素が空気中と水中(淡水)で拡散して移動する距離と時間を示しました。 拡散では拡散係数:Dの値で分子の移動距離が決まります。

このように水中と空気中では酸素分子の移動の早さに1万倍の差があるので、毛細気管内の水分の多寡は昆虫のガス交換にとって重要な問題です。
毛細気管まで拡散した酸素は管壁を通って細胞質や組織内に拡散していきますが、拡散速度は毛細気管の中に比べて1万分の1と小さいので細胞内の流量も減るために、酸素濃度は低下してミトコンドリアに適した1mmHg 程度になっていると考えられます。
また、毛細気管内の酸素消費速度はこの組織への拡散速度で決まるため、安静時での消費は極めて小さくなり、気門の断続的なガス交換サイクル(後述)で酸素は十分に供給できるのでしょう。
この100年間の気管・毛細気管によるガス交換のメカニズムを解明してきた主な研究をまとめてみます。昆虫の呼吸-その3と7とに一部重複しています。
○1919、1920年にKrogh(クロー、1920年ノーベル生理学賞)は気管・毛細気管内の酸素濃度と気管径の測定を行って、大気圧の数%の分圧差があれば気門から細胞まで拡散のみで十分に酸素供給が可能であり腹部の圧迫は気管系の換気にほとんど寄与しないと報告した。(拡散だけでガス交換しているとの説)(Krogh 1920)
○昆虫の呼吸-7で紹介したWeis-Fogh(ヴァイスフォー)は1964年にトンボの胸部では飛翔筋による気管の変形によって十分な換気がされているとの観察を報告し、毛細気管内では大気圧の3.5%以上の分圧差(27mmHg)があれば拡散によるガス交換が適切に行われるとしました。
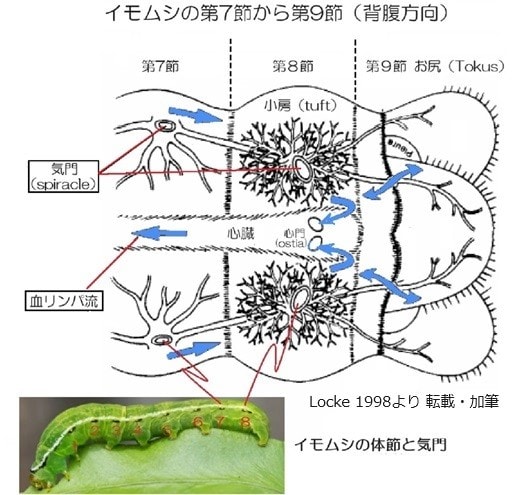
○1998年にLocke、Millはイモムシの第8節の気門に繋がるtuft(タフト、小房)は血リンパ中の血球細胞を酸素化する働きがあり、昆虫の幼虫では気管呼吸以外にそれを補助する酸素吸収機構を持っていると報告した。
○2003年にWestneatは放射光(高速の電子が磁場により方向が帰られるときに電子から放射される光)を用いて、生きている昆虫の気管系の拡張と虚脱による換気運動を撮影し、速い呼吸の時には気管の体積は50%も変化すると報告した(昆虫の呼吸-その3参照)。
○2005年にHetzと Bradleyにより、安静時の昆虫の気門は断続的に開閉して気管内の酸素濃度が約30mmHg(4kPa)に保たれるような不連続なガス交換サイクル(Discontinuous Gas-exchange Cycle:DGC)をしていて、代謝が活発になると連続した換気になると報告した。
DGCにより空気中の150mmHG という高濃度の酸素を30mmHG まで低下させて、酸素によるタンパク質、DNA、脂質の損傷という細胞毒性を避けると同時に、細胞質ではミトコンドリアが効率的に利用する酸素濃度を実現しています。
この様に気管呼吸のメカニズムが理解されてくると、気管は私たち脊椎動物の肺の膨張と収縮に類似した換気運動を行っていることがわかってきました。
これを私たち哺乳類と比べると

となります。昆虫は、は虫類や哺乳類のような大型化への道を進まず、解放血管系+気管呼吸を選択して小型で素早い飛翔という進化を選んだ結果、海中と陸上の全動物種の80%を占める種として繁栄しているのでしょう。
参考文献
1.DL: 2019/2/5
http://www.faculty.ucr.edu insectspagesteachingresourcesstoffolano15.pdf
2. Krogh, A. 1920: Studien uber Tracheenrespiration. ii.
Uber Gasdiffusion in den Tracheen
Archiv fuer die Gesamte Physiologie Berlin 179: 95-112
3.Wigglesworth.V.B. A Theory of Tracheal Respiration in insects
Proceedings of the Royal society B Published:02 April 1930
DL:https://royalsocietypublishing.org/on 16 January 2024
4.その他、昆虫の呼吸-その3と7の文献
前回は細胞に直接空気を供給する気管―毛細気管呼吸を行う昆虫には巨大化に制限がかかるという話でした。細胞数が相似比の3乗に比例して増えること(例えば体長が10倍になれば細胞数は1000倍になること)により気管・毛細気管という空気配管の体積が昆虫体内に占める割合が著しく大きくなることがその要因と考えました。一方、私たち脊椎動物はガス交換のために循環系という血液配管を持つことで巨大化に伴う細胞への酸素供給の問題を解決しています。
今回は気管-毛細気管の構造と水分の移動についてです。
気管は筋肉の収縮や体内の圧力の上昇で圧縮・虚脱し、圧力が弱まるとらせん糸(テニディア)の弾性で拡張するという機械的な運動で換気しています(次図)。一方、毛細気管にはテニディアはなくて直径は1~0.2μmと細く呼吸のためのガス交換は拡散で行われています。(昆虫の呼吸-5、6参照)。その壁の厚さはわずかに0.01~0.03μm(Weis-Foghの原文では100 ~ 300 オングストローム)であり酸素や二酸化炭素の透過にはほとんど影響しないと考えられています。ちなみに酸素分子の大きさは3~4オングストロームです。
この図では直径100μmと150μmの気管と密集して気管を内張しているテニディアが見られます。
毛細気管内の水分移動と酸素拡散
1930年にWigglesworthは蚊の幼虫の観察・実験を基に、毛細気管が細胞内に陥入している抹消では毛細気管内は細胞から浸出した液体で満たされていることを指摘しています。彼の実験では以下のことがわかっています。
・安静時には末梢部分は液体で満たされている(図A)
・窒息や運動中の組織液中には過剰な乳酸がある
・窒息あるいは活動している筋肉では、毛細気管中の液体は吸収されて毛細気管内
の空気が細胞に向かって伸びていく(図B)
・幼虫に塩化ナトリウムと乳酸カリウムを与えて高浸透圧にすると筋肉の活動時と同じ様な液体の吸収と空気柱の伸張がみられる。
これらの結果から、
毛細気管が陥入している細胞質と周囲の組織液は筋肉細胞の活動のために、乳酸やその他の濃度が高くなって、浸透圧が上昇する。
その結果、毛細気管のなかの浸出液は組織中に再吸収されて空気が毛細気管の末梢まで届くようになる。
このように、浸透圧作用により活動時や低酸素時には毛細気管の末梢にまで空気が入って効率よく細胞に酸素が行き渡ると考察しています。
次の表に、酸素が空気中と水中(淡水)で拡散して移動する距離と時間を示しました。 拡散では拡散係数:Dの値で分子の移動距離が決まります。
このように水中と空気中では酸素分子の移動の早さに1万倍の差があるので、毛細気管内の水分の多寡は昆虫のガス交換にとって重要な問題です。
毛細気管まで拡散した酸素は管壁を通って細胞質や組織内に拡散していきますが、拡散速度は毛細気管の中に比べて1万分の1と小さいので細胞内の流量も減るために、酸素濃度は低下してミトコンドリアに適した1mmHg 程度になっていると考えられます。
また、毛細気管内の酸素消費速度はこの組織への拡散速度で決まるため、安静時での消費は極めて小さくなり、気門の断続的なガス交換サイクル(後述)で酸素は十分に供給できるのでしょう。
この100年間の気管・毛細気管によるガス交換のメカニズムを解明してきた主な研究をまとめてみます。昆虫の呼吸-その3と7とに一部重複しています。
○1919、1920年にKrogh(クロー、1920年ノーベル生理学賞)は気管・毛細気管内の酸素濃度と気管径の測定を行って、大気圧の数%の分圧差があれば気門から細胞まで拡散のみで十分に酸素供給が可能であり腹部の圧迫は気管系の換気にほとんど寄与しないと報告した。(拡散だけでガス交換しているとの説)(Krogh 1920)
○昆虫の呼吸-7で紹介したWeis-Fogh(ヴァイスフォー)は1964年にトンボの胸部では飛翔筋による気管の変形によって十分な換気がされているとの観察を報告し、毛細気管内では大気圧の3.5%以上の分圧差(27mmHg)があれば拡散によるガス交換が適切に行われるとしました。
○1998年にLocke、Millはイモムシの第8節の気門に繋がるtuft(タフト、小房)は血リンパ中の血球細胞を酸素化する働きがあり、昆虫の幼虫では気管呼吸以外にそれを補助する酸素吸収機構を持っていると報告した。
○2003年にWestneatは放射光(高速の電子が磁場により方向が帰られるときに電子から放射される光)を用いて、生きている昆虫の気管系の拡張と虚脱による換気運動を撮影し、速い呼吸の時には気管の体積は50%も変化すると報告した(昆虫の呼吸-その3参照)。
○2005年にHetzと Bradleyにより、安静時の昆虫の気門は断続的に開閉して気管内の酸素濃度が約30mmHg(4kPa)に保たれるような不連続なガス交換サイクル(Discontinuous Gas-exchange Cycle:DGC)をしていて、代謝が活発になると連続した換気になると報告した。
DGCにより空気中の150mmHG という高濃度の酸素を30mmHG まで低下させて、酸素によるタンパク質、DNA、脂質の損傷という細胞毒性を避けると同時に、細胞質ではミトコンドリアが効率的に利用する酸素濃度を実現しています。
この様に気管呼吸のメカニズムが理解されてくると、気管は私たち脊椎動物の肺の膨張と収縮に類似した換気運動を行っていることがわかってきました。
これを私たち哺乳類と比べると
となります。昆虫は、は虫類や哺乳類のような大型化への道を進まず、解放血管系+気管呼吸を選択して小型で素早い飛翔という進化を選んだ結果、海中と陸上の全動物種の80%を占める種として繁栄しているのでしょう。
参考文献
1.DL: 2019/2/5
http://www.faculty.ucr.edu insectspagesteachingresourcesstoffolano15.pdf
2. Krogh, A. 1920: Studien uber Tracheenrespiration. ii.
Uber Gasdiffusion in den Tracheen
Archiv fuer die Gesamte Physiologie Berlin 179: 95-112
3.Wigglesworth.V.B. A Theory of Tracheal Respiration in insects
Proceedings of the Royal society B Published:02 April 1930
DL:https://royalsocietypublishing.org/on 16 January 2024
4.その他、昆虫の呼吸-その3と7の文献















※コメント投稿者のブログIDはブログ作成者のみに通知されます