








水中の動物たちの呼吸18―まとめ
水中の動物のシリーズでは魚類と頭足類の呼吸と循環の機能について見てきました。
今回は17回分のまとめです。
原生代末期(先カンブリア時代)のエディアカラ紀から古生代にかけて、海中で発生した大型で扁平な動物たちは、体表から水中の酸素を拡散(酸素が表皮を通して染みこむこと)によって取り込んでいました。口と消化器官を形作り、そこにエラを進化させるまでには更に数千万年の時間が必要でした。
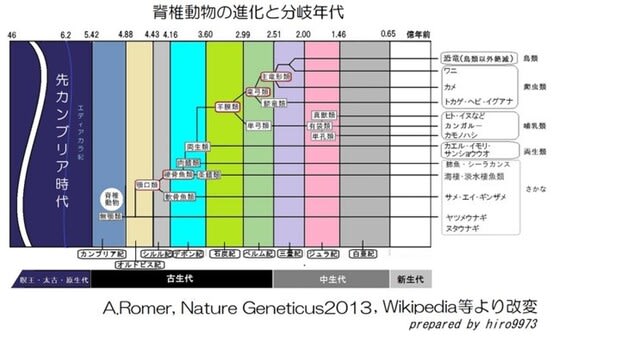
約5.2億年前の古生代カンブリア紀前期には脊椎動物の無顎類(開閉する顎は無くて口は丸く開口)、節足動物(三葉虫など、甲殻類と昆虫の祖先)や軟体動物(オウムガイなど、貝やイカ・タコの祖先)、 半索動物(筆石など、ギボシムシの祖先)が出現しました。
初期の魚類である無顎類では酸素の吸収と二酸化炭素の排出は全身の表皮で行っていました。血液は心臓から全身の組織で酸素を放出し二酸化炭素を取り込んで、皮膚の毛細血管から酸素の吸収と二酸化炭素の排出を行って心臓に戻ります。こうして酸素の豊富な血液がまず始めに心臓に流れていました(血液循環は心臓→全身→皮膚→心臓)。
咽頭にある櫛状の線毛管(エラの元)は口から流入する水から餌をこしとる給餌器官でした。
約4.5億年前のオルドビス紀後期には顎を持つ 魚類(顎口類) が登場し、水流の多い線毛管は給餌器官から酸素を吸収するエラに進化した。
酸素吸収器官が皮膚からエラになると、心臓に流れる血液には酸素が乏しくなり、長時間の高速遊泳や激しい運動時に心臓への酸素供給が不十分になります。
心臓へ還流する静脈血から供給される酸素は冠動脈を持たないスポンジ状心筋にとって必須のものでした。
おそらく、皮膚呼吸に替わる酸素吸収器官が必要になったためでしょうか、硬い骨を持つ魚類はエラを獲得してから約3千万年後のシルル紀中期(約4.2億年前) に肺を進化させました。
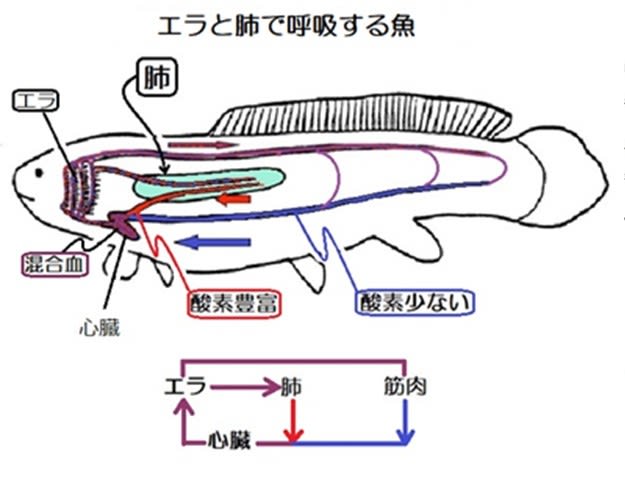
肺で酸素を吸収した血液は心臓へと向かい、全身の組織から帰ってきた静脈血と混合して心臓へと流れていました(血液循環は心臓→エラ→全身→心臓と心臓→肺→心臓とが重複)。
肺を獲得した理由として、「低酸素環境に曝された」ことによる進化圧力との主張が主流ですが、その他に上記のように「心臓へ酸素を供給する」ため、あるいは「浮力を得る」ためなどの生理学的理由も論じられています。
いずれにしろ、エラ呼吸だけでは酸素を十分に取り込むことができない環境の変化に対して、皮膚よりも酸素と二酸化炭素のガス交換に特化した肺を進化させたと言えるでしょう。
ところが、現在のほとんどの魚類は肺を呼吸器官から浮力装置の鰾(ウキブクロ)に変えています。
酸素を吸収する重要な器官である肺をなぜ鰾(ウキブクロ)に変えたのでしょうか。
その理由として
1生存圏を深海に広げたため空気呼吸で水面まで浮上するのが困難になった
2水面で空気を吸うときに翼竜などの空からの捕食者を避けるため
3沈まないための浮力の獲得
4心筋へ酸素を送る効果的な冠状動脈系の発達で低酸素血でも良くなった
などが言われています。
このように魚類の進化では、皮膚 →エラ →エラ+肺 →エラ(+鰾)が主要な呼吸器官の変遷ですが、酸素吸収には肺だけでなく口から消化管の各部位(口腔、のど、食道、胃、腸)までを利用して、それぞれが独立して何度も発生してきました。
例えば現生のトビハゼ(ムツゴロウなど)は口腔内面と皮膚を利用して干潟を這い回っています。木登り魚は上䚡器官(ラビリンス)を呼吸器官に利用し、長時間陸上で過ごすことができます。
トビハゼの皮膚には薄い水の膜があって乾燥を防ぎ、必要な酸素の80%を吸収しています。
湿った皮膚と口腔、咽頭を利用する呼吸方法は両生類が採用している呼吸と同じですが、循環系は基本的には普通の硬骨魚と同じ構造であり、心臓―鰓―全身―心臓と心臓―空気呼吸器官(口腔やラビリンス)―全身―心臓とは並列してつながっています。
肺を使う空気呼吸魚
デボン紀に硬骨魚類から進化・分岐した条鰭類の一部と肉鰭類は現在でも肺を主要な酸素吸収器官としています。特に肉鰭類の肺魚は肺を使って酸素呼吸の90%を肺から採っています。
肉鰭類の心臓は条鰭類と同じ1心房1心室ですが、心房と心室には不完全な中隔があって肺を通って心臓に帰ってきた血液と全身から戻ってきた血流が混合しないように調節されています。このため肺から流出する酸素化された血液は主に全身へと流れ、全身の組織から戻ってきた脱酸素化された血液は肺へと灌流していて、機能的に2心房2心室に類似した血液循環になっています。
肉鰭類から進化した両生類の2心房1心室の心臓では、心室内の弁(らせん弁)により低酸素血は主に肺と皮膚へ流れ、酸素化された血液は大部分が大動脈に流れるという機能的な2心室構造になっていて、肺魚の心臓はこれに類似した血流調整が行われています。
頭足類
古生代カンブリア紀の初期(5.4億年前)に貝殻を持つ最古の頭足類のプレクトロノセラスが現れました。
プレクトロノセラスの系統から、約4.1億年前のシルル紀末期にオウムガイ類があらわれ、更に肉鰭魚類が出現した4億年前のデボン紀には、アンモナイト類とイカやタコの祖先の鞘型類(しょうけいるい)が分岐して出現しました。
タコとイカは、元々は貝殻を作る外とう膜と斜紋筋という筋肉で、鞘(さや)のような中空の筒を作り、その中に内臓を納めているので、これが鞘型類という名前の由来です。
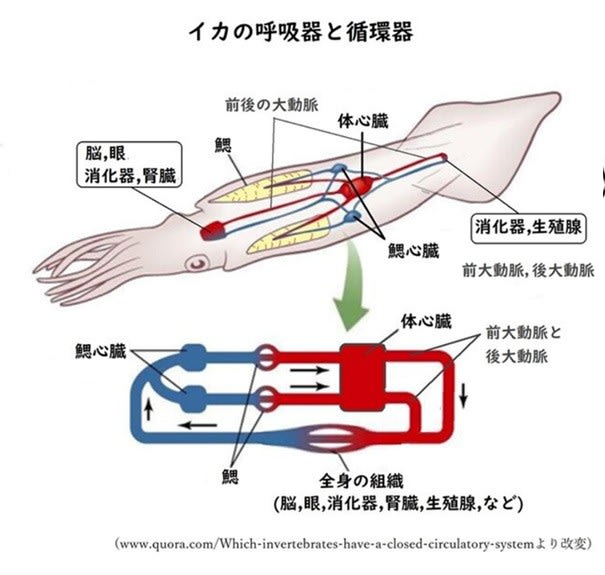
筒の筋肉を使って吸い込んだ海水を左右にある房状のエラに流して酸素を吸収しています。
左右のエラ心臓から押し出された血液はエラを通った後に体心臓に流入して全身の臓器へ循環しています。この心臓の機能を哺乳類と比較すると、右心房と右心室の機能をエラ心臓が担い、左心房左心室を体心臓が受け持っていることになります。ポンプとしての心臓を低圧系のエラ心臓と高圧系の体心臓に分けたために、運動時に必要な酸素を含んだ血液を十分に送り出せるようになっています。
酸素を運ぶ血色素は脊椎動物のヘモグロビン(Hb)ではなくヘモシアニン(Hc)です。
HcはHbより60倍も大きい分子で、酸素運搬の効率はHbよりも少し低く、酸素と結合すると青色になります(脱酸素化すると無色透明になるので、イカ・タコを切っても血は目立ちません)。
古生代初期に、Hcを用いる無脊椎動物が現れ、次いでHbを用いる脊椎動物が出現しました。現在の地球の動物種ではHcを利用する種の方がはるかに多く、昆虫や蜘蛛、エビ、カニなどの節足動物は全動物種の85%を占めて第1位、第2位は貝やタコ、イカなどの軟体動物であり約8%を占め、脊椎動物(魚から鳥、ほ乳類まで)の5%以下よりも遙かに多いのです。
Hc動物のほとんどは解放血管系を持ち小型ですが、タコ・イカだけは閉鎖血管系を持ち大型で高い知能を持っています。閉鎖血管系が知能の発達や大型化をもたらしたのでしょうか。
Hbを採用した魚類は大型化に伴ってシルル紀には肺を獲得してエラと肺を使うようになりましたが、Hcを選択した軟体動物や節足動物たちは肺を必要としませんでした。水中の酸素の吸収と利用にはHcの方がHbよりも有利だったのでしょうか。
イカ・タコはオウムガイから進化したとされていますが、オウムガイと比べるとその身体の構成の独自性(大きな脳、独自の呼吸循環系、高い運動能力、優れた目、洗練された神経系と形態変化能力など)がみられます。これは遺伝子の激変があったためですが、その変化の原因は隕石などからもたらされたレトロウイルスによるというパンスペルミア説(宇宙汎種説)も主張されています。
魚類は肉鰭魚類に進化すると水呼吸から空気呼吸へと切り替えて、陸上へ進出する準備が整いました。いよいよ両生類への進化が始まります。
次回からは魚類や軟体動物と同時期のカンブリア紀前期に出現した三葉虫を代表とする節足動物と、その子孫の昆虫について見ていきます。
その独特の呼吸器官は大変興味深いものです。
参考文献は、「水中の動物たちの呼吸1~17」を参照してください。
水中の動物のシリーズでは魚類と頭足類の呼吸と循環の機能について見てきました。
今回は17回分のまとめです。
原生代末期(先カンブリア時代)のエディアカラ紀から古生代にかけて、海中で発生した大型で扁平な動物たちは、体表から水中の酸素を拡散(酸素が表皮を通して染みこむこと)によって取り込んでいました。口と消化器官を形作り、そこにエラを進化させるまでには更に数千万年の時間が必要でした。
約5.2億年前の古生代カンブリア紀前期には脊椎動物の無顎類(開閉する顎は無くて口は丸く開口)、節足動物(三葉虫など、甲殻類と昆虫の祖先)や軟体動物(オウムガイなど、貝やイカ・タコの祖先)、 半索動物(筆石など、ギボシムシの祖先)が出現しました。
初期の魚類である無顎類では酸素の吸収と二酸化炭素の排出は全身の表皮で行っていました。血液は心臓から全身の組織で酸素を放出し二酸化炭素を取り込んで、皮膚の毛細血管から酸素の吸収と二酸化炭素の排出を行って心臓に戻ります。こうして酸素の豊富な血液がまず始めに心臓に流れていました(血液循環は心臓→全身→皮膚→心臓)。
咽頭にある櫛状の線毛管(エラの元)は口から流入する水から餌をこしとる給餌器官でした。
約4.5億年前のオルドビス紀後期には顎を持つ 魚類(顎口類) が登場し、水流の多い線毛管は給餌器官から酸素を吸収するエラに進化した。
酸素吸収器官が皮膚からエラになると、心臓に流れる血液には酸素が乏しくなり、長時間の高速遊泳や激しい運動時に心臓への酸素供給が不十分になります。
心臓へ還流する静脈血から供給される酸素は冠動脈を持たないスポンジ状心筋にとって必須のものでした。
おそらく、皮膚呼吸に替わる酸素吸収器官が必要になったためでしょうか、硬い骨を持つ魚類はエラを獲得してから約3千万年後のシルル紀中期(約4.2億年前) に肺を進化させました。
肺で酸素を吸収した血液は心臓へと向かい、全身の組織から帰ってきた静脈血と混合して心臓へと流れていました(血液循環は心臓→エラ→全身→心臓と心臓→肺→心臓とが重複)。
肺を獲得した理由として、「低酸素環境に曝された」ことによる進化圧力との主張が主流ですが、その他に上記のように「心臓へ酸素を供給する」ため、あるいは「浮力を得る」ためなどの生理学的理由も論じられています。
いずれにしろ、エラ呼吸だけでは酸素を十分に取り込むことができない環境の変化に対して、皮膚よりも酸素と二酸化炭素のガス交換に特化した肺を進化させたと言えるでしょう。
ところが、現在のほとんどの魚類は肺を呼吸器官から浮力装置の鰾(ウキブクロ)に変えています。
酸素を吸収する重要な器官である肺をなぜ鰾(ウキブクロ)に変えたのでしょうか。
その理由として
1生存圏を深海に広げたため空気呼吸で水面まで浮上するのが困難になった
2水面で空気を吸うときに翼竜などの空からの捕食者を避けるため
3沈まないための浮力の獲得
4心筋へ酸素を送る効果的な冠状動脈系の発達で低酸素血でも良くなった
などが言われています。
このように魚類の進化では、皮膚 →エラ →エラ+肺 →エラ(+鰾)が主要な呼吸器官の変遷ですが、酸素吸収には肺だけでなく口から消化管の各部位(口腔、のど、食道、胃、腸)までを利用して、それぞれが独立して何度も発生してきました。
例えば現生のトビハゼ(ムツゴロウなど)は口腔内面と皮膚を利用して干潟を這い回っています。木登り魚は上䚡器官(ラビリンス)を呼吸器官に利用し、長時間陸上で過ごすことができます。
トビハゼの皮膚には薄い水の膜があって乾燥を防ぎ、必要な酸素の80%を吸収しています。
湿った皮膚と口腔、咽頭を利用する呼吸方法は両生類が採用している呼吸と同じですが、循環系は基本的には普通の硬骨魚と同じ構造であり、心臓―鰓―全身―心臓と心臓―空気呼吸器官(口腔やラビリンス)―全身―心臓とは並列してつながっています。
肺を使う空気呼吸魚
デボン紀に硬骨魚類から進化・分岐した条鰭類の一部と肉鰭類は現在でも肺を主要な酸素吸収器官としています。特に肉鰭類の肺魚は肺を使って酸素呼吸の90%を肺から採っています。
肉鰭類の心臓は条鰭類と同じ1心房1心室ですが、心房と心室には不完全な中隔があって肺を通って心臓に帰ってきた血液と全身から戻ってきた血流が混合しないように調節されています。このため肺から流出する酸素化された血液は主に全身へと流れ、全身の組織から戻ってきた脱酸素化された血液は肺へと灌流していて、機能的に2心房2心室に類似した血液循環になっています。
肉鰭類から進化した両生類の2心房1心室の心臓では、心室内の弁(らせん弁)により低酸素血は主に肺と皮膚へ流れ、酸素化された血液は大部分が大動脈に流れるという機能的な2心室構造になっていて、肺魚の心臓はこれに類似した血流調整が行われています。
頭足類
古生代カンブリア紀の初期(5.4億年前)に貝殻を持つ最古の頭足類のプレクトロノセラスが現れました。
プレクトロノセラスの系統から、約4.1億年前のシルル紀末期にオウムガイ類があらわれ、更に肉鰭魚類が出現した4億年前のデボン紀には、アンモナイト類とイカやタコの祖先の鞘型類(しょうけいるい)が分岐して出現しました。
タコとイカは、元々は貝殻を作る外とう膜と斜紋筋という筋肉で、鞘(さや)のような中空の筒を作り、その中に内臓を納めているので、これが鞘型類という名前の由来です。
筒の筋肉を使って吸い込んだ海水を左右にある房状のエラに流して酸素を吸収しています。
左右のエラ心臓から押し出された血液はエラを通った後に体心臓に流入して全身の臓器へ循環しています。この心臓の機能を哺乳類と比較すると、右心房と右心室の機能をエラ心臓が担い、左心房左心室を体心臓が受け持っていることになります。ポンプとしての心臓を低圧系のエラ心臓と高圧系の体心臓に分けたために、運動時に必要な酸素を含んだ血液を十分に送り出せるようになっています。
酸素を運ぶ血色素は脊椎動物のヘモグロビン(Hb)ではなくヘモシアニン(Hc)です。
HcはHbより60倍も大きい分子で、酸素運搬の効率はHbよりも少し低く、酸素と結合すると青色になります(脱酸素化すると無色透明になるので、イカ・タコを切っても血は目立ちません)。
古生代初期に、Hcを用いる無脊椎動物が現れ、次いでHbを用いる脊椎動物が出現しました。現在の地球の動物種ではHcを利用する種の方がはるかに多く、昆虫や蜘蛛、エビ、カニなどの節足動物は全動物種の85%を占めて第1位、第2位は貝やタコ、イカなどの軟体動物であり約8%を占め、脊椎動物(魚から鳥、ほ乳類まで)の5%以下よりも遙かに多いのです。
Hc動物のほとんどは解放血管系を持ち小型ですが、タコ・イカだけは閉鎖血管系を持ち大型で高い知能を持っています。閉鎖血管系が知能の発達や大型化をもたらしたのでしょうか。
Hbを採用した魚類は大型化に伴ってシルル紀には肺を獲得してエラと肺を使うようになりましたが、Hcを選択した軟体動物や節足動物たちは肺を必要としませんでした。水中の酸素の吸収と利用にはHcの方がHbよりも有利だったのでしょうか。
イカ・タコはオウムガイから進化したとされていますが、オウムガイと比べるとその身体の構成の独自性(大きな脳、独自の呼吸循環系、高い運動能力、優れた目、洗練された神経系と形態変化能力など)がみられます。これは遺伝子の激変があったためですが、その変化の原因は隕石などからもたらされたレトロウイルスによるというパンスペルミア説(宇宙汎種説)も主張されています。
魚類は肉鰭魚類に進化すると水呼吸から空気呼吸へと切り替えて、陸上へ進出する準備が整いました。いよいよ両生類への進化が始まります。
次回からは魚類や軟体動物と同時期のカンブリア紀前期に出現した三葉虫を代表とする節足動物と、その子孫の昆虫について見ていきます。
その独特の呼吸器官は大変興味深いものです。
参考文献は、「水中の動物たちの呼吸1~17」を参照してください。






