昆虫の呼吸-その4
酸素濃度と昆虫のサイズについて
古生代後半には、かもめの大きさのトンボ(メガネウラ:体長30cm、翅の幅75cm)や巨大なカゲロウ(翅の幅48cm)、体長1m体重20kgを超えるサソリなど巨大な節足動物がいたことが化石からわかっています。
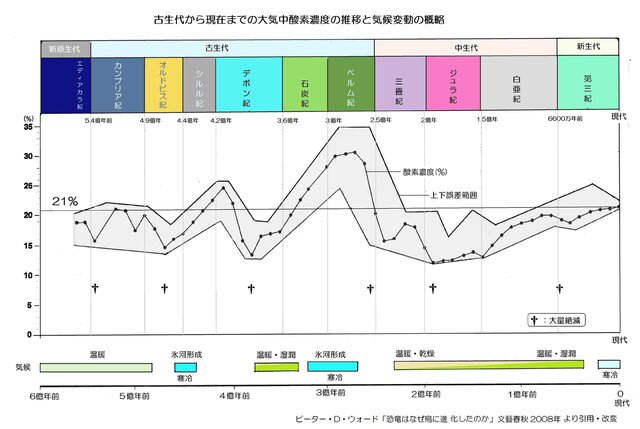
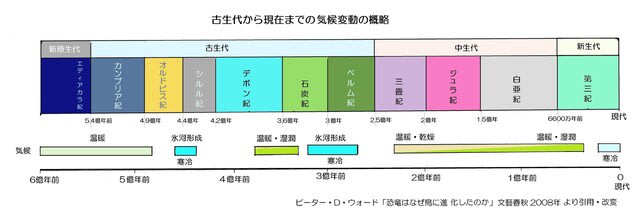
古生代後期の石炭紀からペルム紀にかけての1億年間(3.5~2.5億年前)には酸素濃度が現在の20%よりも高く、最高で35%まで上昇したことが知られています。
・空気中の酸素濃度の変化(古生代から現在まで)
「低酸素の適応―その5」では地球が誕生してから古生代までの40億年間の酸素濃度の変遷を示しましたが、次の図は古生代以後の推移です。
昆虫の巨大化が酸素濃度の高くなった時期と一致しているために、酸素濃度との関連について多くの研究が報告されています。
ピーター・D・ウォードは著書「恐竜はなぜ鳥に進化したのか」の中で酸素濃度が高ければ代謝速度が上がるために巨大化が可能となったとして、酸素濃度の高い条件ではショウジョウバエの体が大きくなる、高濃度の酸素が溶けている水中では水棲節足動物の体が大きい、などの研究を紹介しています(Ward 2008)。
このような酸素濃度の上昇が巨大化の要因とする報告がある一方、酸素濃度が15%前後に下がった中生代白亜紀までは巨大昆虫が発見されているので、必ずしも高酸素濃度が必須であったわけではなさそうです。
酸素濃度と昆虫の大きさについては以下の報告にもみられるように、今でも決着が付いてはいません。
●酸素濃度はサイズの上限を決める部分的要因に過ぎない(Okajima 2008)
●酸素濃度と昆虫のサイズに関連があると考えられるが、多くの要因が関連しているために、古生物学的に詳細なデータと多世代に亘る実験が有用ではないか。(JF Harrison. 2010.)
●幼虫は体表からの拡散で酸素を吸収するために、高酸素濃度による酸素毒性の影響を直接受ける。そのリスクを下げるために、体積当たりの表面積が減るように巨大化したと考えられる。巨大化は成虫ではなく幼虫に原因があるとする報告です。(Verberk 2011)
●巨大化は酸素に関連しているが、その後サイズが小さくなったのは、鳥やコウモリの進化と適応放散のために、サイズが大きいと捕食されやすくなったこと、それと白亜紀大量絶滅で大型昆虫が絶滅したためだろう(Clapham 2012)
この様に、古生代後期の高酸素環境と昆虫の巨大化には関連があると考えられていますが、その理由については統一された見解は無いようです。
しかし、水棲節足動物のエビやカニなどの甲殻類は昆虫と同じように外骨格の脱皮を繰り返して成長しますが、昆虫よりもはるかに大きいサイズになるものがいて、1mを超えるロブスターが捕獲された記録があります。
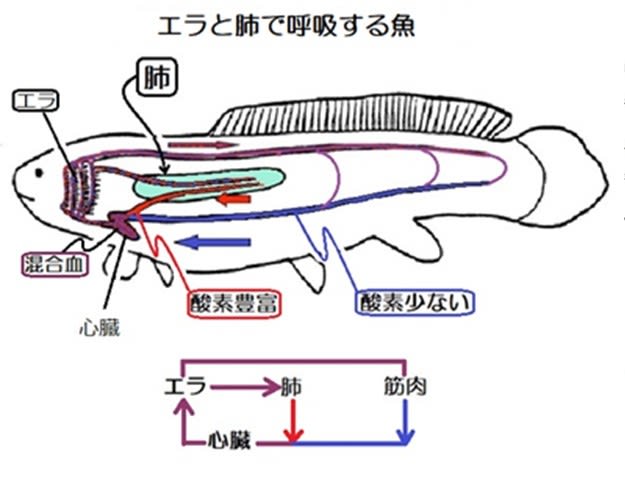
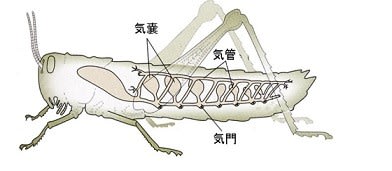
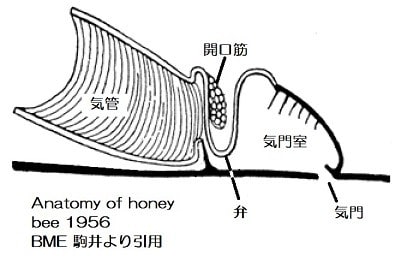
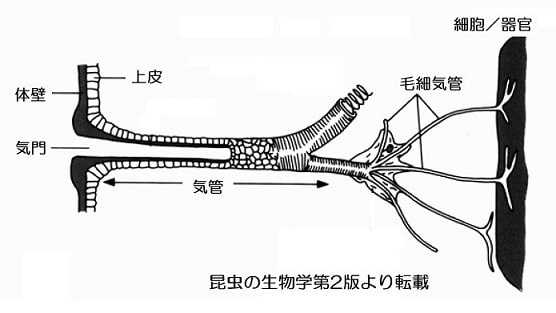
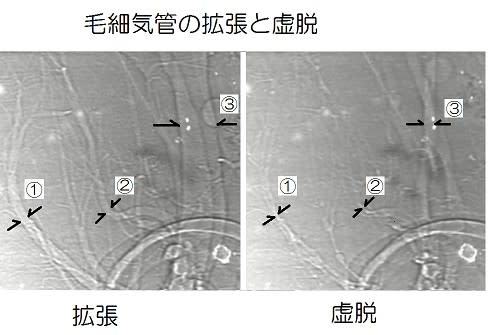
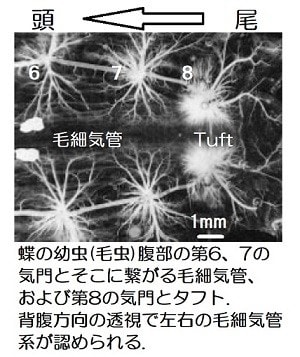
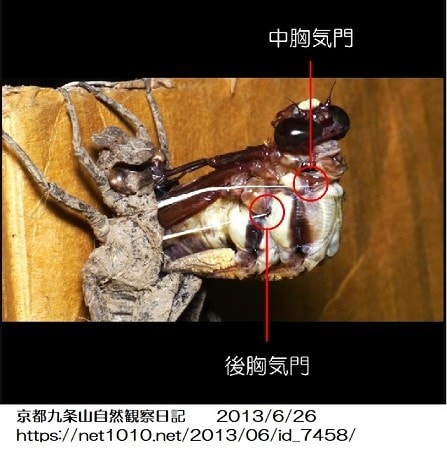
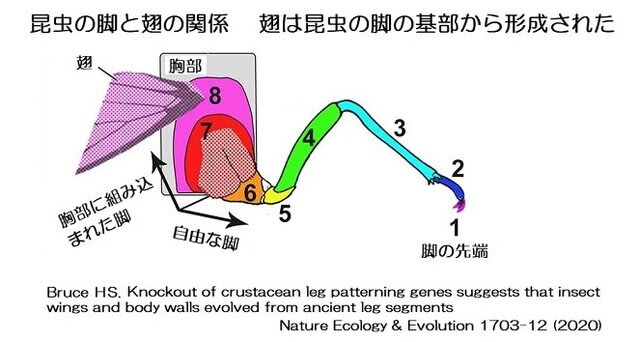
甲殻類と昆虫の大きな差はエラ呼吸と気管呼吸なので、気管-毛細気管系の構造がサイズに関わっているのでしょうか。
参考文献
1.ピーター・D・ウオード 恐竜はなぜ鳥に進化したのか 文藝春秋 2008
2.Okajima, R.: The controlling factors limiting maximum body size of insects. Lethaia2008
3.JF Harrison. Atmospheric oxygen level and the evolution of insect body size. Proceedings of the royal society B. 10 March 2010.
4.Wilco C. E. P. Verberk , David T. Bilton PLoS ONE 6(7): e22610. 2011/7/27 https://doi.org/10.1371/journal
5.Matthew E. Clapham and Jered A. Karr. PNAS July 3, 2012 109 (27) 10927-10930
酸素濃度と昆虫のサイズについて
古生代後半には、かもめの大きさのトンボ(メガネウラ:体長30cm、翅の幅75cm)や巨大なカゲロウ(翅の幅48cm)、体長1m体重20kgを超えるサソリなど巨大な節足動物がいたことが化石からわかっています。
古生代後期の石炭紀からペルム紀にかけての1億年間(3.5~2.5億年前)には酸素濃度が現在の20%よりも高く、最高で35%まで上昇したことが知られています。
・空気中の酸素濃度の変化(古生代から現在まで)
「低酸素の適応―その5」では地球が誕生してから古生代までの40億年間の酸素濃度の変遷を示しましたが、次の図は古生代以後の推移です。
昆虫の巨大化が酸素濃度の高くなった時期と一致しているために、酸素濃度との関連について多くの研究が報告されています。
ピーター・D・ウォードは著書「恐竜はなぜ鳥に進化したのか」の中で酸素濃度が高ければ代謝速度が上がるために巨大化が可能となったとして、酸素濃度の高い条件ではショウジョウバエの体が大きくなる、高濃度の酸素が溶けている水中では水棲節足動物の体が大きい、などの研究を紹介しています(Ward 2008)。
このような酸素濃度の上昇が巨大化の要因とする報告がある一方、酸素濃度が15%前後に下がった中生代白亜紀までは巨大昆虫が発見されているので、必ずしも高酸素濃度が必須であったわけではなさそうです。
酸素濃度と昆虫の大きさについては以下の報告にもみられるように、今でも決着が付いてはいません。
●酸素濃度はサイズの上限を決める部分的要因に過ぎない(Okajima 2008)
●酸素濃度と昆虫のサイズに関連があると考えられるが、多くの要因が関連しているために、古生物学的に詳細なデータと多世代に亘る実験が有用ではないか。(JF Harrison. 2010.)
●幼虫は体表からの拡散で酸素を吸収するために、高酸素濃度による酸素毒性の影響を直接受ける。そのリスクを下げるために、体積当たりの表面積が減るように巨大化したと考えられる。巨大化は成虫ではなく幼虫に原因があるとする報告です。(Verberk 2011)
●巨大化は酸素に関連しているが、その後サイズが小さくなったのは、鳥やコウモリの進化と適応放散のために、サイズが大きいと捕食されやすくなったこと、それと白亜紀大量絶滅で大型昆虫が絶滅したためだろう(Clapham 2012)
この様に、古生代後期の高酸素環境と昆虫の巨大化には関連があると考えられていますが、その理由については統一された見解は無いようです。
しかし、水棲節足動物のエビやカニなどの甲殻類は昆虫と同じように外骨格の脱皮を繰り返して成長しますが、昆虫よりもはるかに大きいサイズになるものがいて、1mを超えるロブスターが捕獲された記録があります。
甲殻類と昆虫の大きな差はエラ呼吸と気管呼吸なので、気管-毛細気管系の構造がサイズに関わっているのでしょうか。
参考文献
1.ピーター・D・ウオード 恐竜はなぜ鳥に進化したのか 文藝春秋 2008
2.Okajima, R.: The controlling factors limiting maximum body size of insects. Lethaia2008
3.JF Harrison. Atmospheric oxygen level and the evolution of insect body size. Proceedings of the royal society B. 10 March 2010.
4.Wilco C. E. P. Verberk , David T. Bilton PLoS ONE 6(7): e22610. 2011/7/27 https://doi.org/10.1371/journal
5.Matthew E. Clapham and Jered A. Karr. PNAS July 3, 2012 109 (27) 10927-10930




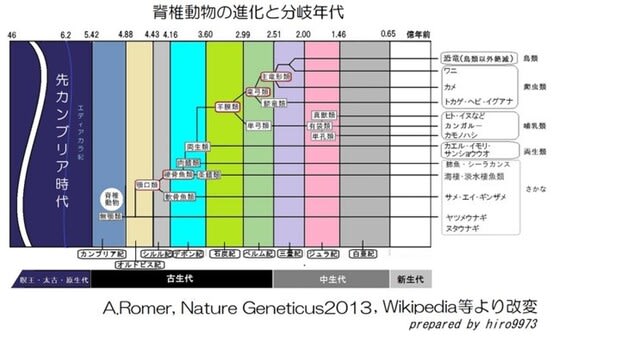
 ・進化史の概略
・進化史の概略