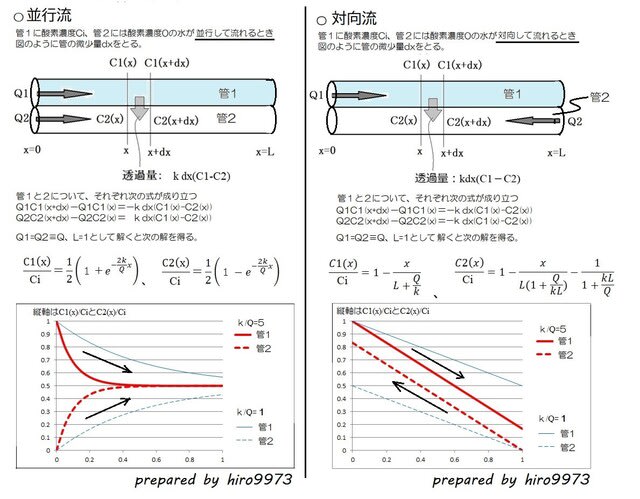
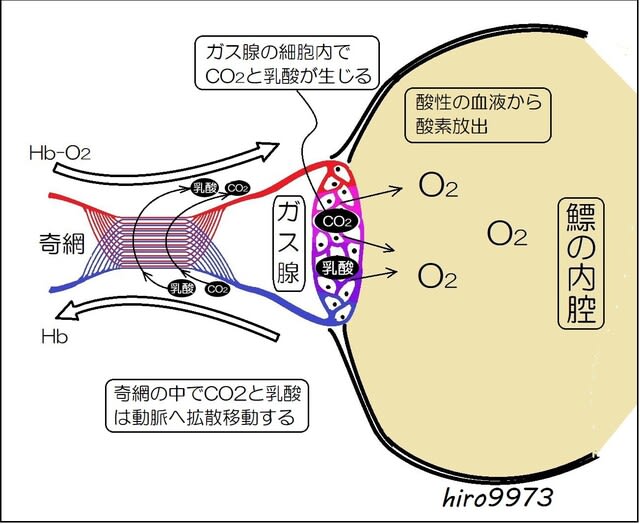
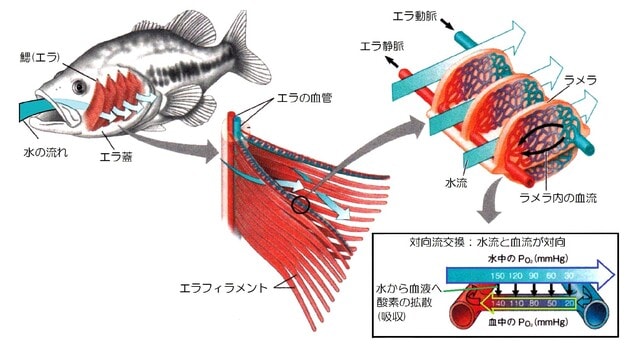
=肺は心臓に酸素を供給するために進化しました!=
原初の魚が肺を獲得した要因についてC. Farmer(以下、ファーマー)は心臓への酸素の供給という仮説を1997年と1999年に報告しました。
その仮説をこの二つの論文から概説します。
たくさんの過去の研究を引用して仮説を少しずつ説明している論文です。
その中で、初期の魚は海で発生し、肺を獲得した環境も従来言われていたような酸素の少ない河川や汽水域ではなく、十分な酸素のある海洋であったと推定しています。
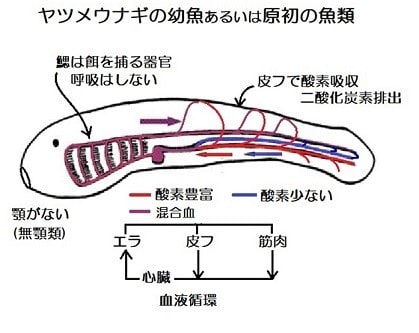
原初の魚
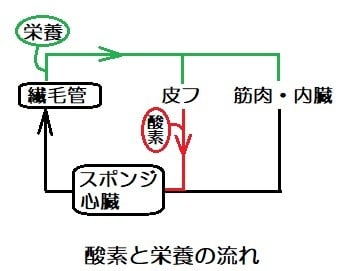
皮フから酸素を十分に吸収した血液は、全身をめぐって酸素が乏しくなった血液と混合されて、心臓に入る。
心臓の筋肉の細胞はスポンジ状であり、心臓の筋肉に血液を送る専用の血管である冠動脈はないので、酸素は柱状の細胞組織の間を流れる血液から供給された。
咽頭の線毛管は、現在の魚の鰓(エラ)のように酸素を吸収する働きはなくて、水中の微少な餌を粘液糸で沪し取り栄養を吸収していた。

(この図は、「硬骨魚18種の心室心筋組織の比較」、 鬼頭・小 栗 (1983)より転載)
エラ呼吸する魚(魚の図はC.ファーマー1997から改変)
皮膚呼吸をやめて、エラだけから酸素を吸収すると、心臓へ流れる血液は酸素が乏しくなる。
活発な遊泳をすると筋肉などで更に酸素が消費されるので、特に運動している時に心臓への酸素はますます足りなくなる。スポンジ状の心筋には冠動脈のように効率的な血管がないことも酸素不足を助長することになる。
更に、低酸素状態では血液は酸性になる(呼吸性アシドーシス)ために、エラで酸素を吸収する能力が低下する(ヘモグロビンに酸素が結合する能力が低下:ボーア効果、ルート効果)。
このように、エラ呼吸のみでは魚が活発な遊泳を行うには不十分でした。
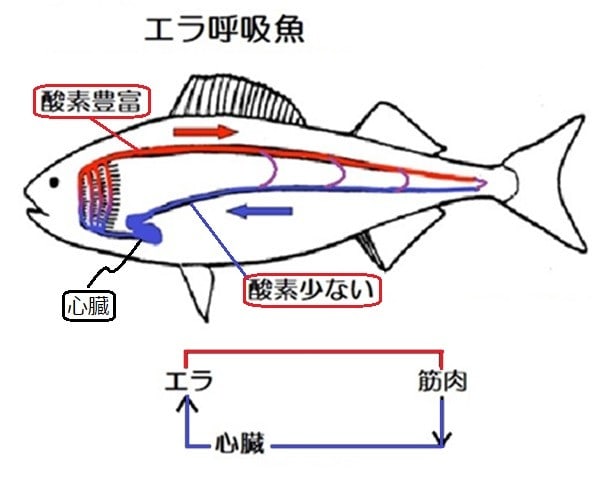
魚にとって心臓に向かう静脈は、心筋細胞に酸素を供給するルートであり、原初の魚が皮膚呼吸から吸収する酸素はスポンジ状心筋にとって必須のものでした。
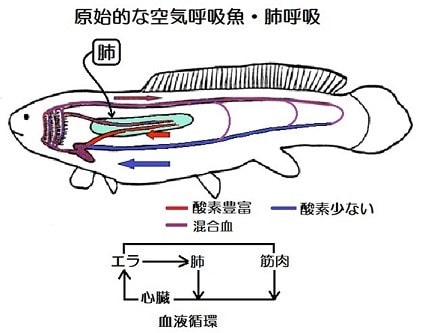
皮膚呼吸の代わりに肺を獲得した魚
エラ呼吸だけでは心臓への酸素を十分に供給できないので、皮膚呼吸の代わりに肺を獲得したと考えられます。
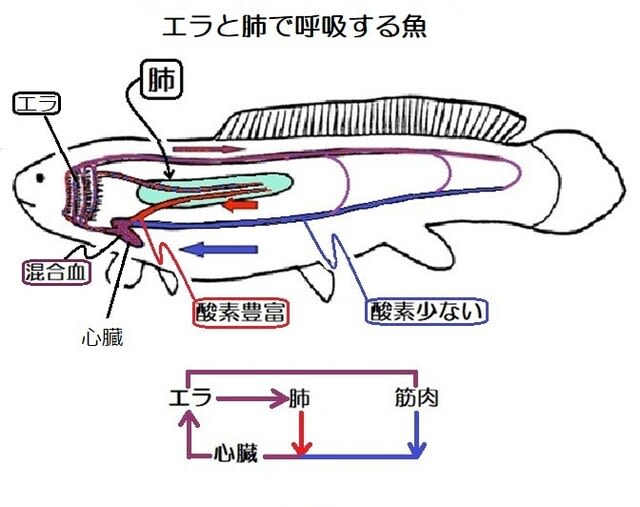
このように、肺を通った血液は酸素を豊富に含んでいるので、心臓の手前で全身から戻ってきた酸素の少ない血液と混合して、その酸素濃度を上げています。この血液がスポンジ状の心筋細胞の間を流れて酸素を供給する。心臓から押し出された血液はエラに流入して酸素を吸収し二酸化炭素を排出したあとに、二つに分かれます。一方は肺へ流れてさらに酸素を吸収して心臓に向かい、もう一方は全身の臓器・筋肉で酸素を放出(二酸化炭素を吸収)して心臓へと向かう。このようなサイクルを繰り返します。
ほ乳類や鳥類では、酸素の豊富な動脈血が心臓にも全身の臓器にも酸素を運ぶ役割を担っていて、静脈血はどの臓器にも酸素は供給していません。その視点から見ると肺からの血液が心臓の手前で静脈に合流する循環は全身への酸素の運搬という点で効率が悪いようにみえますが、スポンジ状の心筋に酸素を送るためには適切な解決法でした。
肺を失った理由について
肺が心臓に酸素を供給する上で重要な原始的臓器ならば、なぜほとんどの現生の硬骨魚類が肺を鰾(ウキブクロ)に変えたのか。その理由として以下が示唆されます。
1生息する水中深度:
肺呼吸をしていた時、生息域は浅い水域に限られますが、深海への進出するに伴い水面まで浮上
して空気を呼吸するのに必要なエネルギーが増加したため肺を喪失した可能性。
2捕食動物:
それまでの捕食者は水生動物だったが、翼竜や鳥類といった空からの捕食者の出現により水面での空気呼吸中の捕食から逃れるために空気呼吸を止めた。
3浮力の獲得
外洋性の魚類では沈まないために使うエネルギーを減らす必要があり、そのために鰾が必要なの
で肺を鰾に変えたとする説(Liem 1988)。鰾を持っていないと沈まないためにエネルギーが必要になる。
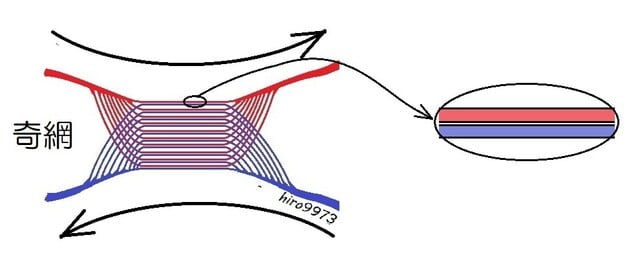
4冠状動脈循環の獲得
スポンジ状心筋に加えて緻密な心筋組織に血液を送り酸素を供給する専用の血管として冠状動
脈が進化・獲得された。肺の代わりに効率のよい血管系を獲得した。
肺の獲得と鰾への進化については概ねこのように概説されています。この肺から鰾への進化は肺を獲得してから硬骨魚類が出現したジュラ紀まで2億年以上かかっているので、ジュラ紀に現れた翼竜や鳥類が主因である可能性が高いようです。
論文ではさらに現生の肺魚の呼吸器と循環器について解説してから、陸に上がった両生類、は虫類の循環系についても検証を進めています。それについてはまた後で。
ふだんは人間や哺乳類の心臓ばかりみているから、魚の心臓の筋肉がスポンジ状だとは思ってもいませんでした。魚の静脈血の酸素化が、冠動脈のないスポンジ状心筋への酸素供給には必須なんですね。人間を基準にして魚や両生類、は虫類の呼吸と循環の効率を判断してはいけないようです。
参考文献
鬼頭・小栗「硬骨魚18種の心室心筋組織の比較」1983
C. Farmer Paleobiology , 1997
C. Farmer Annu. Rev. Physiol. 1999
原初の魚が肺を獲得した要因についてC. Farmer(以下、ファーマー)は心臓への酸素の供給という仮説を1997年と1999年に報告しました。
その仮説をこの二つの論文から概説します。
たくさんの過去の研究を引用して仮説を少しずつ説明している論文です。
その中で、初期の魚は海で発生し、肺を獲得した環境も従来言われていたような酸素の少ない河川や汽水域ではなく、十分な酸素のある海洋であったと推定しています。
原初の魚
皮フから酸素を十分に吸収した血液は、全身をめぐって酸素が乏しくなった血液と混合されて、心臓に入る。
心臓の筋肉の細胞はスポンジ状であり、心臓の筋肉に血液を送る専用の血管である冠動脈はないので、酸素は柱状の細胞組織の間を流れる血液から供給された。
咽頭の線毛管は、現在の魚の鰓(エラ)のように酸素を吸収する働きはなくて、水中の微少な餌を粘液糸で沪し取り栄養を吸収していた。
(この図は、「硬骨魚18種の心室心筋組織の比較」、 鬼頭・小 栗 (1983)より転載)
エラ呼吸する魚(魚の図はC.ファーマー1997から改変)
皮膚呼吸をやめて、エラだけから酸素を吸収すると、心臓へ流れる血液は酸素が乏しくなる。
活発な遊泳をすると筋肉などで更に酸素が消費されるので、特に運動している時に心臓への酸素はますます足りなくなる。スポンジ状の心筋には冠動脈のように効率的な血管がないことも酸素不足を助長することになる。
更に、低酸素状態では血液は酸性になる(呼吸性アシドーシス)ために、エラで酸素を吸収する能力が低下する(ヘモグロビンに酸素が結合する能力が低下:ボーア効果、ルート効果)。
このように、エラ呼吸のみでは魚が活発な遊泳を行うには不十分でした。
魚にとって心臓に向かう静脈は、心筋細胞に酸素を供給するルートであり、原初の魚が皮膚呼吸から吸収する酸素はスポンジ状心筋にとって必須のものでした。
皮膚呼吸の代わりに肺を獲得した魚
エラ呼吸だけでは心臓への酸素を十分に供給できないので、皮膚呼吸の代わりに肺を獲得したと考えられます。
このように、肺を通った血液は酸素を豊富に含んでいるので、心臓の手前で全身から戻ってきた酸素の少ない血液と混合して、その酸素濃度を上げています。この血液がスポンジ状の心筋細胞の間を流れて酸素を供給する。心臓から押し出された血液はエラに流入して酸素を吸収し二酸化炭素を排出したあとに、二つに分かれます。一方は肺へ流れてさらに酸素を吸収して心臓に向かい、もう一方は全身の臓器・筋肉で酸素を放出(二酸化炭素を吸収)して心臓へと向かう。このようなサイクルを繰り返します。
ほ乳類や鳥類では、酸素の豊富な動脈血が心臓にも全身の臓器にも酸素を運ぶ役割を担っていて、静脈血はどの臓器にも酸素は供給していません。その視点から見ると肺からの血液が心臓の手前で静脈に合流する循環は全身への酸素の運搬という点で効率が悪いようにみえますが、スポンジ状の心筋に酸素を送るためには適切な解決法でした。
肺を失った理由について
肺が心臓に酸素を供給する上で重要な原始的臓器ならば、なぜほとんどの現生の硬骨魚類が肺を鰾(ウキブクロ)に変えたのか。その理由として以下が示唆されます。
1生息する水中深度:
肺呼吸をしていた時、生息域は浅い水域に限られますが、深海への進出するに伴い水面まで浮上
して空気を呼吸するのに必要なエネルギーが増加したため肺を喪失した可能性。
2捕食動物:
それまでの捕食者は水生動物だったが、翼竜や鳥類といった空からの捕食者の出現により水面での空気呼吸中の捕食から逃れるために空気呼吸を止めた。
3浮力の獲得
外洋性の魚類では沈まないために使うエネルギーを減らす必要があり、そのために鰾が必要なの
で肺を鰾に変えたとする説(Liem 1988)。鰾を持っていないと沈まないためにエネルギーが必要になる。
4冠状動脈循環の獲得
スポンジ状心筋に加えて緻密な心筋組織に血液を送り酸素を供給する専用の血管として冠状動
脈が進化・獲得された。肺の代わりに効率のよい血管系を獲得した。
肺の獲得と鰾への進化については概ねこのように概説されています。この肺から鰾への進化は肺を獲得してから硬骨魚類が出現したジュラ紀まで2億年以上かかっているので、ジュラ紀に現れた翼竜や鳥類が主因である可能性が高いようです。
論文ではさらに現生の肺魚の呼吸器と循環器について解説してから、陸に上がった両生類、は虫類の循環系についても検証を進めています。それについてはまた後で。
ふだんは人間や哺乳類の心臓ばかりみているから、魚の心臓の筋肉がスポンジ状だとは思ってもいませんでした。魚の静脈血の酸素化が、冠動脈のないスポンジ状心筋への酸素供給には必須なんですね。人間を基準にして魚や両生類、は虫類の呼吸と循環の効率を判断してはいけないようです。
参考文献
鬼頭・小栗「硬骨魚18種の心室心筋組織の比較」1983
C. Farmer Paleobiology , 1997
C. Farmer Annu. Rev. Physiol. 1999