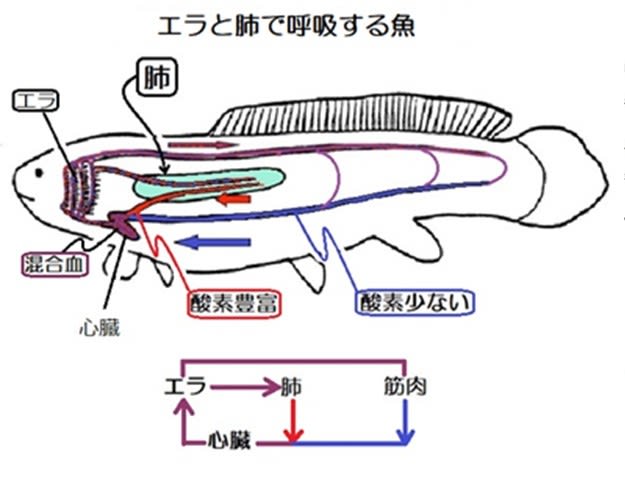
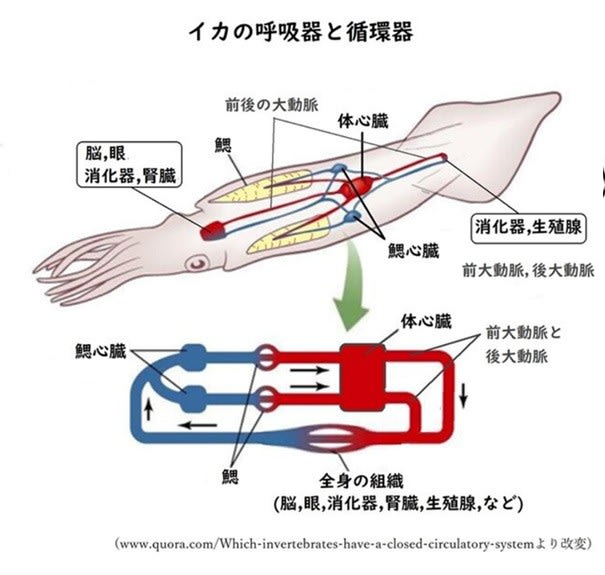
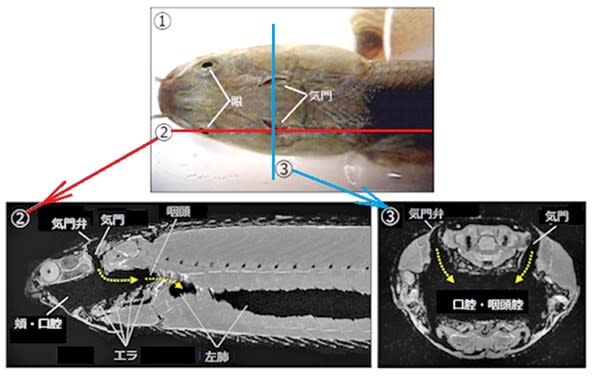
昆虫の呼吸-その3
気管呼吸システムについて:呼吸のための昆虫独自のシステム
昆虫は体表を外骨格で覆っているため体表から酸素を拡散吸収することはできないので、呼吸器官として体表の気門からつながる気管-毛細気管系という独自の気道システムを発達させ、気道内の空気と細胞の間で直接ガス交換を行っています。

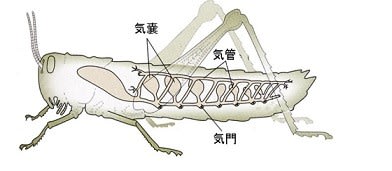
これまでの研究から、気管-毛細気管系における酸素と二酸化炭素のガス交換の様子が明らかになってきました。
気管-毛細気管系の図
図のように外骨格の表面に開口する気門から気管という中空の管が昆虫の体の中を縦横に貫いています。気門は目で見てわかる大きさからダニなどの小型昆虫では数十μm程度で、空気の取り込みと二酸化炭素の排泄時には開き、それ以外は閉じています。
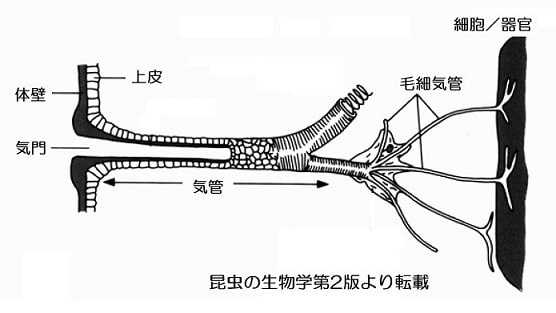
気管は次第に細くなっていき、最終的に直径約0.2μm(1μm=千分の1ミリ)と細い毛細気管となって、各組織中の細胞の間や細胞内に達しています。飛翔筋へは直径0.1μmと更に細くなって細胞内に陥入してミトコンドリアに直接接している場合もあります。
細胞で酸素が消費されると、気門付近の酸素濃度の高い空気から末梢の低酸素の毛細気管内へ酸素が供給されます。その際には幾つかのメカニズムが協同して働いています。
1つは気門から組織までの数mmの毛細気管内を酸素分子が拡散して供給されることです。毛細管内はガス分子の濃度の差により生じる分子拡散現象によって、酸素、二酸化炭素、水の分子が移動しています(拡散流)。酸素は気門から毛細管末梢へ向かい、二酸化炭素と水分子は反対に気門に向かって拡散しています。気管内や組織液中の二酸化炭素は体液中の酵素によって炭酸に変わるために、二酸化炭素分圧は下がっていきます。
拡散流とは:空気の分子は秒速約400mで飛んでいて、分子同士で毎秒数千億回も衝突してランダムに運動しているので濃度が均一に
なる。例えば、酸素ガスの入った箱と窒素ガスの箱を並べて置いて、間の仕切りを取り外すと、この分子同士の衝突によって時間とと
もに酸素と窒素の濃度が均一になる。このようにかき混ぜ無くても均一な濃度になる分子の流れを拡散流という。
2つ目のメカニズムは、飛翔筋など移動に使う筋肉の運動に伴って、気管系が圧迫(虚脱)されて排気し弛緩(拡張)で吸気する換気運動(容積流)です(Weis-Fogh1964)。
2003年には生きている昆虫に放射光といわれる強いX線を照射して直接換気運動をとらえることができました(Westneat 2003)。
図のように気管-毛細気管系は昆虫体の筋肉の収縮と弛緩によって、腹部では背腹方向に虚脱と再拡張が行われている映像が発表されました(インターネット上で閲覧可能です)。
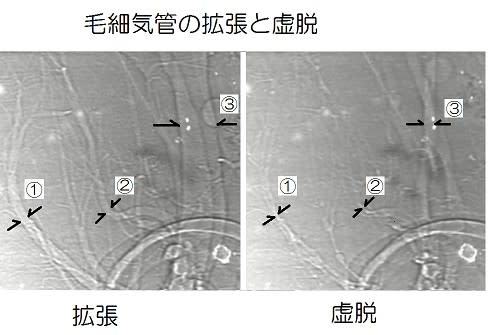
①、②、③の毛細気管は著しく直径が変化しています。さらに細い毛細気管の虚脱も認められます。
虚脱と再拡張により気管内の空気は十分に換気されることになります。
気管から毛細気管まではこの換気運動を用いて空気を入れかえて酸素分圧を上げることで、拡散制限のかかる直径0.2~0.1μmという末梢のガス交換を促していますと思われます(拡散制限は後の回で解説します)。
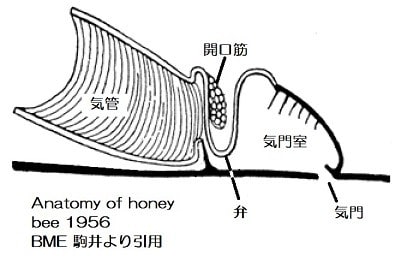
さて、3つ目は気門の働きです。以前から気門の開いている時間は閉じている時間よりもずっと短く、二酸化炭素の排出を調節していることが知られていました。
蛾のさなぎの観察から気門は8時間に一度、約1時間開いて二酸化炭素を排出しています。
酸素が消費されて分圧が下がると気管内の気圧が下がり、気門の隙間から流入する空気で酸素が補給されると考えられてきました。(Miller 1964)
気門の更に積極的な作用が2005年に報告されました。それによると気門は数時間から数日毎に数分間程度開いて断続的なガス交換をします。実験では酸素分圧を0.05~0.5気圧(原文は5~50kPa;100kPaは約1気圧)の間で変化させた環境でも昆虫の気管内酸素分圧は約0.04気圧(4kPa)に保たれていて、高濃度の酸素が組織に障害を起こさない様に、また水分が過剰に蒸散しないように調節されていました。(Hetz 2005.)
なお、4kPaは、約30mmHgに相当します。哺乳類の末梢組織の間質と細胞では酸素分圧は5~40mmHg(平均23mmHg)なので(ガイトン生理学13版p528)、毛細気管内の酸素分圧30mmHgは多くの動物細胞の場合と同程度になっています。
更に呼吸―循環系について1998年に興味深い報告がありました。毛細気管系は直接細胞とガス交換を行うだけでなく、血リンパ中の血球を酸素化していました(Locke 1998)。
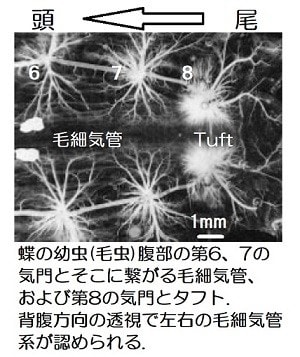
蝶の幼虫(いわゆる毛虫)の背脈管近くの第8気門に繋がる毛細気管末梢にtuft(小房)という構造があります(図)。これは毛細気管が繊細な綿毛のように広がり、きわめて薄い壁で血体腔の血リンパ液中に広がっている構造です。
この小房の周囲には血リンパ中の血球である免疫担当細胞が付着して酸素を受け取っている(酸素化されている)ことがわかりました。つまり小房では、ほ乳類の肺胞で赤血球を酸素化するようにリンパ球を酸素化して、そのリンパ球はすぐ背側にある背脈管(心臓)を通って全身に循環していました。
この様な最近20年間の研究成果から、昆虫の気管-毛細気管系は、単に昆虫の体内に張り巡らされた固定した空気配管ではなく、気門によって酸素濃度を調節し、気管の適切な換気運動を行って組織に直接酸素を供給すると共に循環系の血球を酸素化して全身に送る巧妙な器官です。陸生脊椎動物(両生類、爬虫類、鳥類、ほ乳類)とは全く異なる機構を採用して外骨格と開放血管系に適応して空気呼吸しています。
参考文献
スコット・R・ショー「昆虫は最強の生物である」河出書房新社2016年
松香光夫ほか 昆虫の生物学 第2版 玉川大学出版 1992
Weis-Fogh T. J Exp Biol 41: 229-56, 1964
Westneat,MW. Science vol299 558-560 2003
Miller PL: The Physiology of Insecta Vol. 3, Academic Press, New York, 1964
Hetz, S.K. Nature. 433: 516-519. 2005.
Locke、M。J Insect Physiol. 44(1):1-201998
ガイトン生理学13版p528
Wikipedia 昆虫
気管呼吸システムについて:呼吸のための昆虫独自のシステム
昆虫は体表を外骨格で覆っているため体表から酸素を拡散吸収することはできないので、呼吸器官として体表の気門からつながる気管-毛細気管系という独自の気道システムを発達させ、気道内の空気と細胞の間で直接ガス交換を行っています。
これまでの研究から、気管-毛細気管系における酸素と二酸化炭素のガス交換の様子が明らかになってきました。
気管-毛細気管系の図
図のように外骨格の表面に開口する気門から気管という中空の管が昆虫の体の中を縦横に貫いています。気門は目で見てわかる大きさからダニなどの小型昆虫では数十μm程度で、空気の取り込みと二酸化炭素の排泄時には開き、それ以外は閉じています。
気管は次第に細くなっていき、最終的に直径約0.2μm(1μm=千分の1ミリ)と細い毛細気管となって、各組織中の細胞の間や細胞内に達しています。飛翔筋へは直径0.1μmと更に細くなって細胞内に陥入してミトコンドリアに直接接している場合もあります。
細胞で酸素が消費されると、気門付近の酸素濃度の高い空気から末梢の低酸素の毛細気管内へ酸素が供給されます。その際には幾つかのメカニズムが協同して働いています。
1つは気門から組織までの数mmの毛細気管内を酸素分子が拡散して供給されることです。毛細管内はガス分子の濃度の差により生じる分子拡散現象によって、酸素、二酸化炭素、水の分子が移動しています(拡散流)。酸素は気門から毛細管末梢へ向かい、二酸化炭素と水分子は反対に気門に向かって拡散しています。気管内や組織液中の二酸化炭素は体液中の酵素によって炭酸に変わるために、二酸化炭素分圧は下がっていきます。
拡散流とは:空気の分子は秒速約400mで飛んでいて、分子同士で毎秒数千億回も衝突してランダムに運動しているので濃度が均一に
なる。例えば、酸素ガスの入った箱と窒素ガスの箱を並べて置いて、間の仕切りを取り外すと、この分子同士の衝突によって時間とと
もに酸素と窒素の濃度が均一になる。このようにかき混ぜ無くても均一な濃度になる分子の流れを拡散流という。
2つ目のメカニズムは、飛翔筋など移動に使う筋肉の運動に伴って、気管系が圧迫(虚脱)されて排気し弛緩(拡張)で吸気する換気運動(容積流)です(Weis-Fogh1964)。
2003年には生きている昆虫に放射光といわれる強いX線を照射して直接換気運動をとらえることができました(Westneat 2003)。
図のように気管-毛細気管系は昆虫体の筋肉の収縮と弛緩によって、腹部では背腹方向に虚脱と再拡張が行われている映像が発表されました(インターネット上で閲覧可能です)。
①、②、③の毛細気管は著しく直径が変化しています。さらに細い毛細気管の虚脱も認められます。
虚脱と再拡張により気管内の空気は十分に換気されることになります。
気管から毛細気管まではこの換気運動を用いて空気を入れかえて酸素分圧を上げることで、拡散制限のかかる直径0.2~0.1μmという末梢のガス交換を促していますと思われます(拡散制限は後の回で解説します)。
さて、3つ目は気門の働きです。以前から気門の開いている時間は閉じている時間よりもずっと短く、二酸化炭素の排出を調節していることが知られていました。
蛾のさなぎの観察から気門は8時間に一度、約1時間開いて二酸化炭素を排出しています。
酸素が消費されて分圧が下がると気管内の気圧が下がり、気門の隙間から流入する空気で酸素が補給されると考えられてきました。(Miller 1964)
気門の更に積極的な作用が2005年に報告されました。それによると気門は数時間から数日毎に数分間程度開いて断続的なガス交換をします。実験では酸素分圧を0.05~0.5気圧(原文は5~50kPa;100kPaは約1気圧)の間で変化させた環境でも昆虫の気管内酸素分圧は約0.04気圧(4kPa)に保たれていて、高濃度の酸素が組織に障害を起こさない様に、また水分が過剰に蒸散しないように調節されていました。(Hetz 2005.)
なお、4kPaは、約30mmHgに相当します。哺乳類の末梢組織の間質と細胞では酸素分圧は5~40mmHg(平均23mmHg)なので(ガイトン生理学13版p528)、毛細気管内の酸素分圧30mmHgは多くの動物細胞の場合と同程度になっています。
更に呼吸―循環系について1998年に興味深い報告がありました。毛細気管系は直接細胞とガス交換を行うだけでなく、血リンパ中の血球を酸素化していました(Locke 1998)。
蝶の幼虫(いわゆる毛虫)の背脈管近くの第8気門に繋がる毛細気管末梢にtuft(小房)という構造があります(図)。これは毛細気管が繊細な綿毛のように広がり、きわめて薄い壁で血体腔の血リンパ液中に広がっている構造です。
この小房の周囲には血リンパ中の血球である免疫担当細胞が付着して酸素を受け取っている(酸素化されている)ことがわかりました。つまり小房では、ほ乳類の肺胞で赤血球を酸素化するようにリンパ球を酸素化して、そのリンパ球はすぐ背側にある背脈管(心臓)を通って全身に循環していました。
この様な最近20年間の研究成果から、昆虫の気管-毛細気管系は、単に昆虫の体内に張り巡らされた固定した空気配管ではなく、気門によって酸素濃度を調節し、気管の適切な換気運動を行って組織に直接酸素を供給すると共に循環系の血球を酸素化して全身に送る巧妙な器官です。陸生脊椎動物(両生類、爬虫類、鳥類、ほ乳類)とは全く異なる機構を採用して外骨格と開放血管系に適応して空気呼吸しています。
参考文献
スコット・R・ショー「昆虫は最強の生物である」河出書房新社2016年
松香光夫ほか 昆虫の生物学 第2版 玉川大学出版 1992
Weis-Fogh T. J Exp Biol 41: 229-56, 1964
Westneat,MW. Science vol299 558-560 2003
Miller PL: The Physiology of Insecta Vol. 3, Academic Press, New York, 1964
Hetz, S.K. Nature. 433: 516-519. 2005.
Locke、M。J Insect Physiol. 44(1):1-201998
ガイトン生理学13版p528
Wikipedia 昆虫



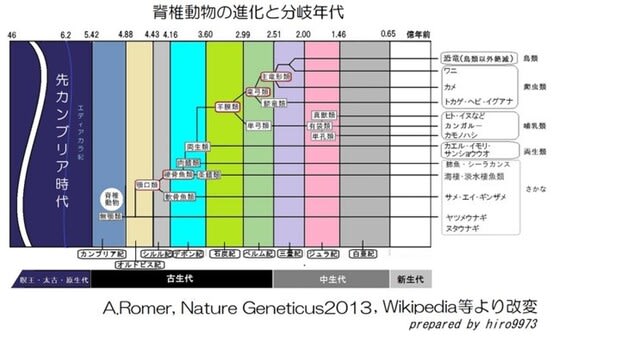
 ・進化史の概略
・進化史の概略