日本人とはどのような人種なのか
後ろに掲載した引用文献からまとめてみました。
遺伝子から見たとき日本列島には絶滅危惧種の遺伝情報が蓄積してきているようです。これまでの研究で分りつつある所です。
完全に理解しているわけではないので生煮えで済みません、もし参考になればということです。
日本人の遺伝子は古くて新しい
日本人は、はるか昔のネアンデルタール人の血さえも濃厚に混ざった世界最古の超古代人種(C1a1系統5%・D1b系統35%/縄文系40%*)と、最先端人種(O1b2系統30%・O2系統20%/弥生系50~55%**)のハイブリッドだったのです。
このC系統は、現在、C1a1とC2を合わせて、日本人の8.3~10%を占める。進取の気性に富み、非常に活動範囲が広く、楽天的で行動である。〔A?CT?CF?C〕
このC1a1の親戚のC1a2は、現代でもヨーロッパに極めて少数存在し、同時にスペインの7000年前の人骨から発見されています。そこから、C1aは、6万年前に出アフリカを果たした、もっとも初期のクロマニヨン人の血統ではないか、とも言われているのです。
現存系統の拡散開始は約12,000年前であり、縄文時代の開始とほぼ一致している。すなわちハプログループC1a1は日本に縄文文化をもたらした集団かもしれない。
7300年前、鬼界カルデラの超巨大噴火が起きた。
鬼界カルデラは屋久島の北にある。 25×15 ㎞の大きさで、薩摩硫黄島や竹島が外輪山である。この噴火は日本列島で最も新しい超巨大噴火である。噴出量170 ㎞3以上でアカホヤ噴火ともよばれる。火砕流は海上を走り、屋久島、種子島、九州南部を覆った。火山灰は東北南部まで達し、紀伊や山陽で20㎝を超えたと推定されている。宮崎県南部では60㎝を超えたとされている。
この火砕流で、南九州に住んでいた縄文人は壊滅したようである。そこに住んでいた縄文人の一部は生き延びて他の地域に移り住んだようである。それは、南九州の先進文化が2000~4000年を経て本州に伝えられていることからも解る。そして、以後1000年間人が住んだ形跡が見られない。その後に住み着いた前期縄文人の系統(D1a2a)は今までの系統(C1a1)と異なるようである。
Y染色体ハプログループC1a1の人は日本最古の住民と言うことである。何をして生計を維持したかは解らないが子孫は数万年を経て現存している。
旧石器時代人の遺伝子としてハプログループD1a2a (Y染色体)が想定されている[4]。
日本固有のD1a2a系統はその発生から3.5-3.7万年ほどたっているとされ[22]、考古学から求められる日本列島に最初に現生人類集団が到来した時期と一致している。
このハプログループD1a2aはアイヌ人の75%に見られることから、D系統はかつての縄文人(旧石器時代のシベリア)のものであると考えられている。但し縄文人のハプログループがD1a2aだけだった訳ではなくハプログループC1a1とハプログループKとハプログループFも縄文人由来と考えられている[15][16]。
縄文人の主要な遺伝子として、ハプログループD1a2a (Y染色体)、ハプログループC1a1 (Y染色体)が想定されている[4]。
父系遺伝子Y染色体ハプログループD1a2aを持つのは世界で本土日本、アイヌ民族、沖縄だけであり、お隣の台湾や中国、韓国には全く見られない。
世界的にも珍しいY染色体ハプログループD1a2a系統を日本人では30%~40%、アイヌでは90%、沖縄本島では50%もの人が所持していることも重要である。
アイヌ人の父系系譜を示すY染色体ハプログループの構成比については、日本列島固有のハプログループ D1a2a が 87.5%(うちD1a2a*が13/16=81.25%、D1a2a1aが1/6=6.25%)と大半を占める。
ハプログループ D1a2aは日本列島以外ではほぼ確認されず、縄文人特有の系統であったと考えられている。これは琉球人で50%弱、本土日本人で30%ほどであるため、アイヌ人は現代日本人の中では縄文人の遺伝子を最も色濃く引き継いでいると言える。
D1b、C1a1と並んで、このO1b2a1aもまた、日本人固有の遺伝子と言ってよいと思います。ちなみに朝鮮半島南部には、弟筋のO1b2a1bが、33%存在します。おそらく、日本から渡ったO1b2a1が、半島で変異したのでしょう。つまり、稲作文化は、順番としては日本列島から朝鮮半島に渡ったようにも見えるのです。
引用ーーーーーー
3.C(第三世代)?およそ7万年前に、エチオピアでAから分離し発生したCTは、ユーラシアン・アダムと呼ばれる。さらに、このCTが、6.5万年前に、アフリカ北東部でCFとDEに分離した。その後、6万年前に、おそらくはアフリカ北東部か西アジアで、CFはCとFに分離し、その両方が出アフリカを果たした。
このうち、特に、C系統は、〝冒険遺伝子〟とも呼ばれ、最初に出アフリカを果たし、世界中に広がった。このC系統のうち、最古の系統C1a1は、現在、日本のみに2.3~4.5%(125~250万人)存在する。沖縄(6%)に多く、アイヌには存在しないので、南方系の海洋航海民の縄文遺伝子とも考えられている。ただし、徳島(10%)や青森(7%)でも、沖縄以上に頻度が高いので、やはり北方系ではないか、という意見もある。
このC1a1と最も近縁のC1a2は、ヨーロッパに、ごくわずかに見られる。このC1a2は、旧石器時代人の人骨や新石器時代人(7000年前のスペインの人骨)からも発見されており、最古のクロマニヨン人の系統に繋がるのではないか、とも考えられている。
なお、出アフリカ後、C1a系統は、およそ5万年前に、C1a2が西(欧州)へ、C1a1は東(東アジア)へと移動した。しかし、C1a1の日本までの移動ルートは、C1a1が日本にしか現存しないため、まったくの謎である。
また、近似種のC1bのうち、C1b1は西アジアに残り、C1b2は南方(オセアニア)へ向かった。そして、C1b2は、5万年前に、ニューギニア島とオーストラリアに達した。現在、クック諸島に82%、アボリジニ・サモア・タヒチに65%、マオリに43%、トンガに35%、フィジーに22%存在する。このC1b2系統は、南太平洋のオセアニア地域一帯に広がっているということだ。
その一方で、C2は、3万年前、ユーラシア大陸内陸部の中央アジアで派生し、現存するハプログループとしては、最初にアメリカ大陸に渡り、南アメリカ大陸の最南端にまで達した。現在、カザフ人に67%、モンゴル人に55%、満州族に27%、ウズベク人に20%、シャイアン・アパッチなどアメリカインディアンに15%、アイヌ・朝鮮に12%、漢民族・日本人全体では5.5~6%ほど存在する。このC2は、ユーラシア大陸の北方遊牧民の遺伝子と考えられている。チンギス・ハーン、ヌルハチ、堺正章はC2である。
このC系統は、現在、C1a1とC2を合わせて、日本人の8.3~10%を占める。進取の気性に富み、非常に活動範囲が広く、楽天的で行動的である。〔A?CT?CF?C〕
ーーーーーーー
ハプログループDのうち、日本でD1a2、チベットでD1a1が高頻度であり、両者は四万年乃至五万年前にわかれた[24][25]。また、O1b-M268には下位分類O-M95とO1b2-M176が存在し、それらの最も近い共通祖先も約三万年前にさかのぼると推定されている。ハプログループCにも日本固有のC1a1-M8とユーラシアに広く分布するC2-M217が含まれるが、両者の最も近い共通祖先は約五万年前と推定されている。C2-M217だけを取ってみても、日本ではC-F1067(日本及び朝鮮、中国等に於けるC-M217の多数を占める)とC-L1373(中央アジア、モンゴル以北の北東アジア、並びにアメリカ大陸先住民に於けるC-M217の多数を占める)の両方が観察されており、両者の最も近い共通祖先は三万年乃至四万年前にさかのぼると推定されている。
ーーーーーーー
約4万年前…
Y染色体C1系統(縄文系)
約4万年前の旧石器時代に、無人の日本列島に初めてやって来た人々。
東南アジアからスンダランドを北上してやって来た海洋民族で、浅瀬の海で漁撈生活をしていたと考えられている。
のちにやって来たハプログループD系統の人々と混ざり、縄文文化を形成した。
沖縄県具志頭村から出土した古代人骨・港川人(約1万7000年前)は、オーストラリア先住民と骨格が近似しているため、Y染色体はハプログループC系統のC1a1(C-M8)に連なる系統にあたると推定。
ーーーーーーー
3万5千年前…
おそらく、ネアンデルタール人と共存・交配したのは、Y染色体ハプログループで言えば、7万年前に〝出アフリカ〟したユーラシアン・アダム(CT)から、およそ7~6万年前に分岐したC系統とD系統とF系統と考えられます。そのどれもが、日本の縄文系・弥生系に含まれますし、特に日本固有種であるC1a1やD1bは、共に「アジア人ですらない」とまで言われるユーラシア大陸最古の種族ですから、約6万年前に中東地域で起こったとされるネアンデルタール人との最初の交雑の該当種の直接の子孫と考えて、何ら不思議ではないどころか、極めて自然であると考えられます。
また、ルーマニアの洞窟から発見された3万2000年前の頭骨のいくつかには、ネアンデルタール人と現生人類の両者の特徴が認められるという研究発表もあります。ホモ・サピエンス・サピエンスとネアンデルタール人との交雑は、数万年に渡って、かなり頻繁に起こっていたのかもしれません。
そして、彼らC1a1やD1bもまた、繰り返される交雑の結果として生まれたネアンデルタール人とのハーフやクォーターらとともに、東へ向けて大陸横断の旅を続け、長い時間をかけて、およそ4~3万年前に、この極東の地、当時、まだ大陸と地続きだった日本にまでやってきたのでしょう。
日本人は、はるか昔のネアンデルタール人の血さえも濃厚に混ざった世界最古の超古代人種(C1a1系統5%・D1b系統35%/縄文系40%*)と、最先端人種(O1b2系統30%・O2系統20%/弥生系50~55%**)のハイブリッドだったのです。
ーーーーーーー
ハプログループC1a1(M8)は日本列島固有であり、その祖型の移動ルートは謎に包まれている。最も近縁なC1a2(V20)は、旧石器時代のチェコ、約7000年前の中石器時代のスペイン北西部および同じ頃の新石器時代のハンガリー、そして少数の現代ヨーロッパ人、カビル人、アルメニア人、ネパール人から検出されており、ヨーロッパ最古層(クロマニョン人)の集団にもそのハプログループに属す者がいた。もう少し遡ると、インドやアラビアなどに散見されるC1b1(M356)、インドネシア東部からメラネシアおよびポリネシア、オーストラリアの先住民アボリジニに多く見られるC1b2(B477)との共通祖先C1にたどり着く。崎谷満はC1a1の祖型はイラン付近からアルタイ山脈付近を経由し朝鮮半島経由で日本に到達したとしている[8]。その渡来年代は定かでないが、現存系統の拡散開始は約12,000年前であり、縄文時代の開始とほぼ一致している。すなわちハプログループC1a1は日本に縄文文化をもたらした集団かもしれない。https: //ja.wikipedia.org/wiki/ハプログループC1a1_(Y染色体)拡散開始は約12,000年前
ーーーーーーー
DNA解析により、日本人は主にY染色体ハプログループD1a2aの縄文系とハプログループO1b2の弥生系を起源とする事が明らかになった。このハプログループD1a2aはアイヌ人・沖縄人・本土日本人の3集団に多く見られるタイプであり、朝鮮半島や中国人(漢民族)には全く見られない。このハプログループD1a2aはアイヌ人の88%に見られることから、D系統はかつての縄文人(旧石器時代のシベリア)のものであると考えられている。但し縄文人のハプログループがD1a2aだけだった訳ではなくハプログループC1a1も縄文人由来と考えられている[12]。
ーーーーーーー
縄文文化は、根菜・雑穀の単純農耕除き、ほとんど農耕がない採集狩猟の生活でありながら、一年を通じて定住するという、世界でも珍しいタイプの生活・文化圏です。青森県の三内丸山遺跡のように、縄文中期(BC3000年頃/5000年前)に、およそ1500年間にわたって、最盛期には500人の定住者がいたと推測されている遺跡もあります。
そして、この時期に、大陸にはまったく存在しない縄文特有のもう一つの父系遺伝子型(C1a1:4.2万年前に発生?)がすでに存在したと考えられます。ちなみに、このC1a1は、現在、日本人に2.3~4.5%ほど存在し、日本以外の地域ではまったく見られません。
沖縄(6.6%)に多く、アイヌ(0%)にいないため、東南アジア由来の南方系海洋縄文人の血統とも考えられています。しかし、沖縄以上に、むしろ青森(7.7%)や徳島(10%)にも多いので、南方系と断言はできません。また、このC1a1の親戚のC1a2は、現代でもヨーロッパに極めて少数存在し、同時にスペインの7000年前の人骨から発見されています。そこから、C1aは、6万年前に出アフリカを果たした、もっとも初期のクロマニヨン人の血統ではないか、とも言われているのです。
その後、およそ5万年前に、C1a1は東に、C1a2は西へ分かれたのです。ただし、C1a1が、どうやって日本にたどり着いたのか、その経路は完全に闇に閉ざされています。北方からという説もあれば、南方からという説もあります。
ただ、遠い親戚であるC1b2は、海洋狩猟民で、5万年前に、すでにオーストラリアに到達しています。ですから、C1a1の日本への経路も、南からだろうという意見が強いのです。とは言え、遠い親戚ですから、決定的な根拠にはなりません。ともかく、経路はどうあれ、このC1a1は、D1bが日本に到達するよりはるか以前、4.5万年前に日本列島にたどり着いた、最古の日本人かもしれないと言われています。
さらに、2万年前には、北方からC2系統の北方遊牧民が多少入ってきたようです。こちらは、現在、モンゴル人に55%、朝鮮人で10%、日本人では6%が保有し、アイヌでは12%と多く、沖縄では0%です。この分布が、C2が北方から来たとされる根拠です。
現在、日本人のY染色体ハプログループの割合は、O系統(弥生系)が50%、D系統が40%、C系統が8.3%です。O系統は弥生人ですから、縄文人のY染色体は、D1bが主流で、それに、すでに日本列島にいたC1a1、さらに北方からC2が流入して、多少混ざったものだったと考えられます。特にD1bとC1a1は、どちらも非常に古いY染色体の遺伝子配列で、しかも、日本以外では、ほとんど見られないものです。
さて、その後、長江文明の衰退と春秋戦国時代(BC770年~)の戦乱によって、中国南方系の人々(初期弥生人/新モンゴロイド/O1b系統)が、紀元前10世紀ぐらいからポツリポツリと日本列島に渡ってくるようになりました。彼ら、初期弥生人が保有していた父系遺伝子配列が、O1b2と考えられています。この遺伝子の一つに、日本にしか存在しない派生遺伝子O1b2a1aがあります。
彼ら初期弥生人(O1b2系統)は、長江文明にルーツを持ち、黄河文明に駆逐されて、江南から山東や日本列島へと移動した人々と考えられています。主に九州や本州西部に移住し、稲作と金属器を日本に持ち込みました。大陸からきた弥生人は比較的少数で、平野部分や盆地に定住し、土着の縄文人の方は、主に海岸や山地で生活していたので、当時は、ある程度の住み分けと交流、および共生が可能だったようです。
ちなみに、この〝弥生遺伝子〟の主流であるO1b2a1aは、4000年ほど前に長江流域で発生したO1b2を祖型とし、日本で派生したものです。現在、日本の全国平均で25%を占めますが、沖縄では11%、アイヌでは0%です。その他の国では、インドネシアで8%、朝鮮で4%、ベトナムで3%ほど見られますが、ほかではほとんど見られません。中国本土でも、O1b2は、戦闘的な漢民族(O2)によって駆逐され、今では影も形もないのです。D1b、C1a1と並んで、このO1b2a1aもまた、日本人固有の遺伝子と言ってよいと思います。ちなみに朝鮮半島南部には、弟筋のO1b2a1bが、33%存在します。おそらく、日本から渡ったO1b2a1が、半島で変異したのでしょう。つまり、稲作文化は、順番としては日本列島から朝鮮半島に渡ったようにも見えるのです。
ーーーーーーー
海洋民族
Y染色体ハプログループC1a1系統 【日本固有種・縄文系】. 浅瀬の海で漁撈生活をしていた海洋民族で、 約4万2,000年前 の旧石器時代に、東南アジアからスンダランドを北上して、無人の日本列島に初めてやって来た人々であると考えられている。
沖縄県具志頭村・港川1号人骨(港川人)
沖縄県具志頭村から出土した古代人骨・港川人(約1万7000年前)は、オーストラリア先住民と骨格が近似しているため、Y染色体はハプログループC系統のC1a1(C-M8)(注1)に連なる系統にあたると推定されている(注2)。
遼寧省錦州の男性
中国・遼寧省錦州大業郷出身の男性から得られたY染色体を解析した結果、日本固有種と同じハプログループC1a1(C-M8)が検出された(注1)。
Y染色体ハプログループC1b2系統
アボリジニー
オーストラリア大陸とタスマニア島などの原住民族である・アボリジニー(Aborigine)のY染色体は、ハプログループC1b2b(C-M347)が60%、パプア・ニューギニアなどに多い系統のハプログループK2(K-M526)が22%である(注1)。
ーーーーーーー
D系統のなかでも「D1b(旧D2)」は日本人しかもっておらず、特に「アイヌ人」に9割近くも集積し、沖縄本島にも多いです。
D1b: 日本(大和)36% (琉球)45% (アイヌ)88%
C3 :12%
毛深い、色白、ホリが深いという白人的な特徴をもつ「アイヌ人は縄文人の子孫」であると言われます。
日本人のY染色体の割合
縄文系:D1b(D-M64.1) 36%、C1a1(C-M8) 5%
弥生系:O1b2a1a1(O-CTS10145) 25%、
O1b(O-M268) 10%
渡来系:O2(O-M122) 20%
に大別されます。
ーーーーーーー
C1a1系統はパプア系に違和感があります
C系統は現生人類の中で最初に世界中に拡散したハプロです。その中で日本人に多いC系統はC1a1系とC2系の2系統があります。C2系はツングース系に多いハプロですから問題はないと思います。但し、C2系統の下位区分ではツングース系民族に多いC2aではなく、C2bが日本人を含めた東アジア人には多いようです。
C1a1については、パプアニューギニア人やアボリジニに多いC1b系ではなく、ヨーロッパ人の最古層のクロマニョン人のハプロであり、現代ヨーロッパ人にも少数だが存在するC1a系から分岐したハプロです。縄文人のハプロにもいた可能性もありますが、おそらくは日本に渡来した時期は弥生系の人々と一緒にきたと思われます。その痕跡が中国人や韓国人にもこのハプロがいることが判明しています。
ーーーーーーー
DNA解析により、日本人は主にY染色体ハプログループD1a2aの縄文系とハプログループO1b2の弥生系を起源とする事が明らかになった。このハプログループD1a2aはアイヌ人・沖縄人・本土日本人の3集団に多く見られるタイプであり、朝鮮人には少数みられるものの、中国人(漢民族)にはまず見られない。このハプログループD1a2aはアイヌ人の75%に見られることから、D系統はかつての縄文人(旧石器時代のシベリア)のものであると考えられている。但し縄文人のハプログループがD1a2aだけだった訳ではなくハプログループC1a1とハプログループKとハプログループFも縄文人由来と考えられている[15][16]。
ハプログループD系統はYAP型(YAPハプロタイプ)ともいわれ、現代アジアにおいて支配的なO系統やC2系統とは分岐から7万年以上経ており、最も近縁であり同じYAP型であるE系統とも6.5万年前に分岐した系統である。
現在D系統は、日本列島以外で高頻度の地域はチベット[17][18]とアンダマン諸島[19]しかない。
ハプログループDは、現代の東アジア人とシベリア人に近い「東アジアのハイランダーズ」(古代チベット人)に関連していて、縄文時代後期に増加し、C1a1、K、Pなどの以前の縄文時代の系統に大きく取って代わったとされる[20][21]。
なお、当時の弥生人や現代東アジアにおいて支配的なO系統は、ウラル系のN系統やコーカソイド系において最多的なR系統などと近縁であり、YAP型(D系統、E系統)とは全く異なるグループである。以上のことから縄文人から自然進化的に弥生人が派生したという説は完全に否定されている。
++++そうなのか
日本列島にD系統の人々が入ってきたのは数万年前の最終氷期地続きの時代と考えられている。その証拠として、日本人のD系統にのみ見られる多くのSNPの発生があげられる。SNPは突然変異により発生する確率的な事象であるから、発生数によって時間の経過が分かるのである。日本固有のD1a2a系統はその発生から3.5-3.7万年ほどたっているとされ[22]、考古学から求められる日本列島に最初に現生人類集団が到来した時期と一致している。
++++そうなのか
長らく縄文人の人骨よりY染色体ハプログループは分析されてこなかったが、北海道礼文島の船泊遺跡(縄文時代後期前葉から中葉(約3,800~3,500年前))から出土した人骨・船泊5号のY染色体ハプログループがD1a2a2a(D-CTS220)であることが判明した[23]。これにより「ハプログループD1a2aは縄文系である」という従来よりの仮説に一つ近づいたが、検証したのは長い縄文時代の中期のたった一人である為、時代、地域共に広範囲での多数の検証が期待される。とりわけハプログループC1a1は拡散年代と縄文文化開始の時期が一致しており、今後の研究いかんによっては初期の縄文人の主要なDNAとなる可能性がある。C1a1とC2aは日本人固有であり、現在の日本ではおよそ5%の頻度で発見されている
ーーーーーーー
分岐年代でわかるY染色体ハプログループChttp://digx.hatenablog.com/entry/y-C2tree
「日本のC1a1の拡がった時期」とか、「オーストラリアにはアボリジニ(C1b2b-M347)が早い時期にたどり着いていた」とか。
C2(旧C3)が増加し始めたのはC1より全体的に遅く、C2b・C2c1のレベルでも最終氷期の末期。C2cのみ共通祖先で34500という数字が出てる。
ただこれは、この最終氷期末期に何らかの理由でボトルネックが生じた、と見るべきか。詳細は後で。
C1b1-K281は中国からも発見されている。人口の多いインドなど中心でそれなりに多いこともあり、人口を計算すると、日本のC1a1よりもずっと多い集団となる模様。
C2は大まかに、北方のC2b系統と南方のC2c系統に分かれる。
ーーーーーーー
日本には縄文文化以前に「旧石器時代の最終に現れた細石器(細石刃)を使った細石刃文化」があった。本州でこの文化のもっとも古い年代は静岡県休場(やすみば)遺跡で、C14年代測定法によれば14,300年前で、終末は12,000年前と言う。大きく分けると、日本列島の細石刃文化は、北東日本の楔形細石刃と南西日本の野岳・休場型や船野型細石刃の二つの分布圏に分かれる。前者はシベリアから北海道を経由して本州へ、後者は中国黄河中・下流から九州を経由して本州へ及んだらしい。日本は細石刃文化(2300年間)、縄文文化(1万年間)、弥生文化(600年間)が古代の三主要文化であり、担い手も違う人々だったようである。
細石刃文化はY-DNA C(5%)、縄文文化はY-DNA D(35%)、弥生文化はY-DNA O(50%)(構成比は現在の各説を平均したもの。正確ではない)となっており、一万年の長きにわたった縄文文化は北は北海道から南は沖縄まで日本全国あまねく行き渡り、地域間における多様性も豊かだったようである。
細石刃文化は今は滅んだ文化なのではあるが、2000年の長きにものを言わせ担い手は現在も滅んではいない。弥生文化はある意味現在も継続しており、Y-DNA Oは増える一方だ。私見で恐縮だが、これは戦後のベビーブームが影響しており、江戸時代まではY-DNA Dが圧倒的に多かったのではないか。
Y-DNA Cは子系統にC1とC2があるそうで、C1a1(C-M8)は日本固有と言い、C1a2(C-V20/V184)およびその子系統はヨーロッパに多いので、人類の出アフリカ後、中央アジアあたりで東西に分岐したと言うことか。C2はベトナムを除いては中央アジア、北東アジア、北アメリカの寒冷地に多いようである。
その役割としては、C1はもっぱらユーラシア大陸の東の果てを目指してやって来て、当時は大陸の一部だったと思われる現在の北海道に到達したのではないか。言わば、Y染色体ハプログループC1a1の人は日本最古の住民と言うことである。何をして生計を維持したかは解らないが子孫は数万年を経て現存している。
C2は、いわゆる、マンモスハンターとしてシベリアから北海道へやって来たのではないか。日本にはマンモスの遺物は北海道にしかないという。津軽海峡(ブラキストン線)を渡るのは大形のマンモスには無理で小形の人間だけが渡ったと言うことか。
また、C1については(Hammer et al.2006)の調査によると九州地域のC1が「0<ゼロ>」という。これはとりもなおさず九州地域のC1の由来(他の調査では少ないながらも九州にもC1の人はいる)は朝鮮半島ではなく本州からやって来たと言うべきか。
C2についても細石刃文化の担い手というものの日本の細石刃文化はバイカル湖発祥の北海道・本州のものと黄河流域発祥の九州の二種類がある。いずれの担い手もY-DNA C2だったのだろうか。
ーーーーーーー
M7a系統は縄文人骨全体に多いが、北海道から東北の縄文人骨はN9b系統が圧倒的に多くなっています。南ルートではM7a系統、北ルートではN9b系統が渡来してきたことを示しているようです。現代日本人(アイヌや沖縄人を含めて)においても、この2つのハプロはそこそこ存在しています。またこの2つ以外のハプロも縄文人骨には見受けられるから、様々な人々が日本列島にやってきて交雑したようです。おそらくYハプロも、Q、N、C1a1、C2、P(QとRの祖型)あたりもいたと思います。
縄文人はMTハプロ分析においても、おそらくは北方のツングース系民族と南方のホアビン文化人系統の交雑民族です。
ーーーーーーー
Y染色体ハプログループC1a1系統 【日本固有種・縄文系】. 浅瀬の海で漁撈生活をしていた海洋民族で、 約4万2,000年前 の旧石器時代に、東南アジアからスンダランドを北上して、無人の日本列島に初めてやって来た人々であると考えられている。http: //famousdna.wiki.fc2.com/wiki/Y%E6%9F%93%E8%89%B2%E4%BD%93C1%E7%B3%BB%E7%B5%B1
ーーーーーーー
浅瀬の海で漁撈生活をしていた海洋民族で、約4万2,000年前の旧石器時代に、東南アジアからスンダランドを北上して、無人の日本列島に初めてやって来た人々であると考えられている。のちにやって来たハプログループD系統の人々と混ざり、縄文文化を形成した。このC1a1系統は、YAP因子を持たない縄文人の系統である。
Y染色体ハプログループC1b1系統
コステンキ遺跡出土人骨
1954年に、ロシアヴォロネジ州のドン川近くにあるコステンキ遺跡から発掘された旧石器時代(約3万8,700-3万6,200年前)の狩猟採集者である若い男性の古人骨・コステンキ14号のY染色体は、ハプログループC1b1*(C-K281*)である(注1)(注2)。
Y染色体ハプログループC1b2系統
アボリジニー
オーストラリア大陸とタスマニア島などの原住民族である・アボリジニー(Aborigine)のY染色体は、ハプログループC1b2b(C-M347)が60%、パプア・ニューギニアなどに多い系統のハプログループK2(K-M526)が22%である(注1)。
沖縄県具志頭村・港川1号人骨(港川人)
沖縄県具志頭村から出土した古代人骨・港川人(約1万7000年前)は、オーストラリア先住民と骨格が近似しているため、Y染色体はハプログループC系統のC1a1(C-M8)(注1)に連なる系統にあたると推定されている(注2)。
ーーーーーーー
Y染色体C1系統
Y染色体ハプログループC1a1系統 【日本固有種・縄文系】
浅瀬の海で漁撈生活をしていた海洋民族で、約4万2,000年前の旧石器時代に、東南アジアからスンダランドを北上して、無人の日本列島に初めてやって来た人々であると考えられている。のちにやって来たハプログループD系統の人々と混ざり、縄文文化を形成した。このC1a1系統は、YAP因子を持たない縄文人の系統である。
ーーーーーーー
ハプログループCにも日本固有のC1a1-M8とユーラシアに広く分布するC2-M217が含まれるが、両者の最も近い共通祖先は約五万年前と推定されている。C2-M217だけを取ってみても、日本ではC-F1067(日本及び朝鮮、中国等でのC-M217の多数を占める)とC-L1373(中央アジア、モンゴル以北の北東アジア、並びにアメリカ大陸先住民でのC-M217の多数を占める)の両方が観察されており、両者の最も近い共通祖先は三万年から四万年前にさかのぼると推定されている。
ーーーーーーー
D1b、C1a1と並んで、このO1b2a1aもまた、日本人固有の遺伝子と言ってよいと思います。
ちなみに朝鮮半島南部には、弟筋のO1b2a1bが、33%存在します。おそらく、日本から渡ったO1b2a1が、半島で変異したのでしょう。
つまり、稲作文化は、順番としては日本列島から朝鮮半島に渡ったようにも見えるのです。
ーーーーーーー
まず、最初に、日本人のY染色体の遺伝子配列(ハプログループ)には、世界中でも日本にしか見られない配列が、三種類もあるということに注目したいと思います。
一つは前述のD1b(35~40%)であり、もう一つはC1a1(2.3~4.5%)、それからO1b2a1a(22~25%)があります。さらに、この三種類のハプログループは、遡れば、出アフリカを果たした三つの系統、CとDEとFに、それぞれ繋がります。
このように、出アフリカを果たした全系統に渡って、三種類ともが、国内にほぼ均等に存在する地域は、世界でも珍しいのです。しかも、それぞれのハプログループが、その地域だけにしか現存していないとなると、そんな希少な地域や国は他に見当たりません。
こうした日本人固有の世界に類のない独特の父系DNA組成は、現在のアイヌ人・琉球人に繋がる古モンゴロイド種の縄文人と、紀元前5世紀以降に中国大陸から渡ってきた新モンゴロイド種の弥生人との間での、平和的な共生と強制的でない緩やかな混血によって生まれました。
縄文人が、今日のアイヌ・沖縄の人々との関係が深いことは、すでに1980年代からの研究で、M7aという母系遺伝子の配列(ミトコンドリアDNAハプログループ/mtDNA)が、日本人(7%)の中でもアイヌ(16%)と沖縄(23%)に特に多く、縄文人の骨からも複数検出されていることから、以前から注目されていました。M7aは、25000年ほど前に、M7から分岐した母系グループで、現在、日本にもっとも多く、特に沖縄とアイヌに多い母系遺伝子です。他では、朝鮮半島で3%ほど見られます。このM7aを持つ人々は、南方から来たのではないか、という説が、定説でした。つまり、この説によれば、縄文人は南から来たということになります。
その後、1990年代以降、DNA解析技術の急速な進歩によって、父系遺伝子の配列が明らかになってきました。そして、前述のように、日本人にしか見られない特殊な父系の遺伝子組成の存在が、注目されています。
この縄文人DNA(ハプログループD1b)は、日本人全体の35~40%(本土では東北・関東で高く、九州北部・四国・中国で低い)、沖縄では45~56%(中でも本島北部と宮古島は特に高い/石垣・八重山諸島では4%しかない)、アイヌ人では88%が保有しています。
ところが、その他の民族では、朝鮮民族でわずかに4.0%、ミクロネシアで5.9%いるだけで、台湾にも中国にも満州にもモンゴルにもチベットにも東南アジア諸国にも一切保有者がいないのです。日本の周辺諸国でさえ、このDNA配列を共有する民族は、ほぼ皆無ということです。
ただ、チベットには、親戚筋に当たるハプログループD1aを持つ人が48%います。そして、日本とチベット以外に、D系統のY染色体配列を10%以上保有する民族・地域は、現在、地球上にほぼ存在しません。
このD系統の遺伝子は、北方系古モンゴロイドの遺伝子と考えられており、日本列島は、チベットとともに、古モンゴロイドの血統を受け継ぐ希少な地域なのです。古モンゴロイドは、およそ6万年前に、アフリカ北東部で黒人種(E系統)と分化した最も古い黄色人種の種族で、後に、2.5万年前に、北東アジアで極北遺伝子Nと分岐したO系統の新モンゴロイドが大陸に分布した後も、険しいチベット高原と海を隔てた日本列島にだけ残ったものと思われます。ただ、もっとも近い親戚関係にあるチベット人(D1a)と日本人(D1b)も、4万年前に分岐したと考えられています。その後、3.5万年前のマンモスハンターとして、D1b種族は氷河を越えて、当時は大陸と地続きだった日本列島に、北方からやってきたのかもしれません。少なくとも、D1bが、南方系(スンダランド由来)という確たる証拠はないのです。
ですから、日本人は、朝鮮人とも中国人ともモンゴル人とも異なる、固有の遺伝配列を持つ、北方系ないしは南方系の古代人種の末裔なのです。また、日本人全体の中でも、原日本人である縄文人の遺伝子(D1b)を最も多く持つのがアイヌ人(88%)であり、そのアイヌ人に一番近いのが、本土の和人ではなくて、距離の遠く離れた沖縄の宮古島であるということです。
特に、血液中で白血球が抗体を作ってきた歴史を辿ることによって、人種間の混交の仕方を血液型から判別できるGm遺伝子配列を見ると、アイヌ人と宮古人との近似性が非常に高いようです。実際、宮古島は、沖縄本島とも石垣島(八重山諸島)とも、言葉も容貌も異なります。彫りの深い顔立ちは、アイヌと宮古に共通する北方系の特徴です。私見ですが、宮古とアイヌは、伝統的な民謡や舞踊にも、どこか共鳴するものを感じます。
宮古島には山がなく、水源に乏しいので、沖縄本島や石垣島と異なり、寒冷な蝦夷地(アイヌの土地)と同様に稲作が不可能でした。それゆえ、大陸から渡ってきた弥生人(新モンゴロイド)が定住することがなく、混血がすすまなかったのでしょう。そのため、アイヌと宮古だけに、色濃く縄文の血統と文化が保全されたものと思われます。
ーーーーーーー
古モンゴロイドの一派である縄文人(後期旧石器時代人)は、第三氷河期の終わりにあたる1.6万年前、太平洋の水位の上昇によって、それまでユーラシア大陸と地続きだった日本列島が、日本海を挟んで大陸から分離して以来、およそ1.3万年(BC16000年~BC1000年)に渡って独自の新石器文化を築いていきます。北は北海道から南は沖縄まで、一つの閉ざされた生活圏・文化圏が形成されたのです。
縄文文化は、根菜・雑穀の単純農耕除き、ほとんど農耕がない採集狩猟の生活でありながら、一年を通じて定住するという、世界でも珍しいタイプの生活・文化圏です。青森県の三内丸山遺跡のように、縄文中期(BC3000年頃/5000年前)に、およそ1500年間にわたって、最盛期には500人の定住者がいたと推測されている遺跡もあります。
そして、この時期に、大陸にはまったく存在しない縄文特有のもう一つの父系遺伝子型(C1a1:4.2万年前に発生?)がすでに存在したと考えられます。ちなみに、このC1a1は、現在、日本人に2.3~4.5%ほど存在し、日本以外の地域ではまったく見られません。
沖縄(6.6%)に多く、アイヌ(0%)にいないため、東南アジア由来の南方系海洋縄文人の血統とも考えられています。しかし、沖縄以上に、むしろ青森(7.7%)や徳島(10%)にも多いので、南方系と断言はできません。また、このC1a1の親戚のC1a2は、現代でもヨーロッパに極めて少数存在し、同時にスペインの7000年前の人骨から発見されています。そこから、C1aは、6万年前に出アフリカを果たした、もっとも初期のクロマニヨン人の血統ではないか、とも言われているのです。
その後、およそ5万年前に、C1a1は東に、C1a2は西へ分かれたのです。ただし、C1a1が、どうやって日本にたどり着いたのか、その経路は完全に闇に閉ざされています。北方からという説もあれば、南方からという説もあります。
ただ、遠い親戚であるC1b2は、海洋狩猟民で、5万年前に、すでにオーストラリアに到達しています。ですから、C1a1の日本への経路も、南からだろうという意見が強いのです。とは言え、遠い親戚ですから、決定的な根拠にはなりません。ともかく、経路はどうあれ、このC1a1は、D1bが日本に到達するよりはるか以前、4.5万年前に日本列島にたどり着いた、最古の日本人かもしれないと言われています。
さらに、2万年前には、北方からC2系統の北方遊牧民が多少入ってきたようです。こちらは、現在、モンゴル人に55%、朝鮮人で10%、日本人では6%が保有し、アイヌでは12%と多く、沖縄では0%です。この分布が、C2が北方から来たとされる根拠です。
現在、日本人のY染色体ハプログループの割合は、O系統(弥生系)が50%、D系統が40%、C系統が8.3%です。O系統は弥生人ですから、縄文人のY染色体は、D1bが主流で、それに、すでに日本列島にいたC1a1、さらに北方からC2が流入して、多少混ざったものだったと考えられます。特にD1bとC1a1は、どちらも非常に古いY染色体の遺伝子配列で、しかも、日本以外では、ほとんど見られないものです。
さて、その後、長江文明の衰退と春秋戦国時代(BC770年~)の戦乱によって、中国南方系の人々(初期弥生人/新モンゴロイド/O1b系統)が、紀元前10世紀ぐらいからポツリポツリと日本列島に渡ってくるようになりました。彼ら、初期弥生人が保有していた父系遺伝子配列が、O1b2と考えられています。この遺伝子の一つに、日本にしか存在しない派生遺伝子O1b2a1aがあります。
彼ら初期弥生人(O1b2系統)は、長江文明にルーツを持ち、黄河文明に駆逐されて、江南から山東や日本列島へと移動した人々と考えられています。主に九州や本州西部に移住し、稲作と金属器を日本に持ち込みました。大陸からきた弥生人は比較的少数で、平野部分や盆地に定住し、土着の縄文人の方は、主に海岸や山地で生活していたので、当時は、ある程度の住み分けと交流、および共生が可能だったようです。
ちなみに、この〝弥生遺伝子〟の主流であるO1b2a1aは、4000年ほど前に長江流域で発生したO1b2を祖型とし、日本で派生したものです。現在、日本の全国平均で25%を占めますが、沖縄では11%、アイヌでは0%です。その他の国では、インドネシアで8%、朝鮮で4%、ベトナムで3%ほど見られますが、ほかではほとんど見られません。中国本土でも、O1b2は、戦闘的な漢民族(O2)によって駆逐され、今では影も形もないのです。D1b、C1a1と並んで、このO1b2a1aもまた、日本人固有の遺伝子と言ってよいと思います。ちなみに朝鮮半島南部には、弟筋のO1b2a1bが、33%存在します。おそらく、日本から渡ったO1b2a1が、半島で変異したのでしょう。つまり、稲作文化は、順番としては日本列島から朝鮮半島に渡ったようにも見えるのです。
ーーーーーーー
例えば、美しい豊穣の女神だったメデューサが、蛇の髪をした人を石に変える恐ろしい化け物にされてしまったように、世界の他の地域では、母系の神々は、すべて男性神によって打ち倒されてしまいました。日本だけが、この相対立する神々を共存させることに成功しているのです。
また、日本の主婦は、夫の収入を完全に管理していて、夫にお小遣いを渡している人が多いのですが、これはアメリカ人などからすると考えられないことです。アメリカ人の夫ならば、自分の給料は完全に自己管理するのが当たり前で、奥さんには生活費とお小遣いしか渡しません。
こうした日本人の独自性を生み出した源は、わたしたちの身体に色濃く流れる縄文の血です。実際、完全に縄文人のDNA配列を受け継いでいるのが、日本人の約5割(D1b?35~40%、C1a1・C2?8~10%)に過ぎないとしても、日本人の9割は、縄文人の遺伝子配列(血)の影響を、大なり小なり何らかの形で、受けていると言われています。その影響が色濃いか薄いかの違いはあるとしても、やはり、古来からの伝統を受け継ぐ日本人であることに違いはないのです。
ーーーーーーー
それから、はっきりさせておきたいことは、「日本人(日本民族)とは何か?」を考える上で、アイヌ人と沖縄人は、決して除外すべき異民族ではないということです。
それどころか、アイヌと沖縄(特に宮古島)にこそ、日本人のルーツがあると言っても過言ではないのです。ですから、2008年に国連が、沖縄を日本の中の少数民族地域と勧告したことは、明らかに誤りです。アイヌ人のY染色体ハプログループは、縄文系(D1a)88%、縄文系北方遊牧民(C2)12%で、圧倒的に縄文人です。沖縄人の場合も、縄文系(D1a・C1a1)61%、弥生系(O1b2・O2)39%で、縄文系が濃厚です。
ですから、ここ千数百年の歴史の歩みの中で、不幸にも一時的に、はぐれてしまったことがあったとは言っても、1万年の日本民族の血脈の中では、アイヌ人も沖縄人も、同じ一つの日本民族であることに違いはないのです。
そう考えると、沖縄の古神道や精霊信仰(アニミズム)や先祖崇拝には、仏教や儒教を受容する以前の日本人の信仰心のあり方が、色濃く反映されており、沖縄人の精神世界には、縄文人の〝心〟が宿っているように思われるのです。アイヌの人々の心のあり方についても、同様のことが感じられます。
伝統的な土着の精神文化に支えられた個人の内面は、世界と運命への信頼に基づく静かで安定した安らぎの中にあります。彼らの心は、疑いとざわめきの中で生きる現代人の心とは対極にあるものです。なぜなら、そのような土着の精神文化を否定し、侮蔑し、消し去ろうとするところから、近代の教育はスタートしたからです。現代文明自体が、固有の精神文化を解体する装置として機能しているのです。そのため、高い教育を受けた者ほど、そのような土着の精神文化を卑下する文明の罠に深く囚われています。
〝現代人の心の再生〟をテーマとする時、わたしたちは、このことをよくよく考えてみなければなりません。
繰り返しますが、日本人の遺伝子の半分は、ユーラシア大陸で最古の遺伝子(C1a1・D1b)であり、残りの半分は最も新しい種類の遺伝子(O1b2・O2)なのです。このようなバランスを持つ地域は、地球上でも本当に稀です。
そのせいか、この国には、最も古いものが、最も新しいものと無理なく自然に共存するという、世界に類のない特性が、至る所で見られます。融和と共生の精神が息づいています。世界の絶滅危惧種であるD系統ハプログループが、この国にこれほど多く生き残っていることの意味を、深く考えてみてください。
この国に深く根付いているポリシーは、「決して敵を滅ぼさない」「根絶やしにしない」ということです。「敵を滅ぼす者は、自らも滅ぼされる。」わたしたちの祖先は、その真理を歴史からも学び、その知恵を「平家物語」「太平記」のようなカタチでも語り継いできました。
たとえ、歴史上、不幸な時期がいくつかあったとしても、そのことをもって、この国の本質を見誤ってはなりません。先の大戦の不幸は、日本人の心に、いまだ大きな影を投げかけてはいますが、それでもあえて言わせていただきたいのです。「歴史の真実をみつめましょう」と。
「平和を尊び、神と人を信頼する人々が、絶えることなく数万年を生きてきた国。」それが、私たちの住む国、日本です。
ーーーーーーー
ハプログループC1a1(M8)…縄文時代に日本から移住…
ハプログループC1a1(M8)の祖型の移動ルートは謎に包まれている。最も近縁なC1a2(V20)は、旧石器時代のチェコ、約7000年前の中石器時代のスペイン北西部および同じ頃の新石器時代のハンガリー、そして少数の現代ヨーロッパ人、カビル人、アルメニア人、ネパール人から検出されており、ヨーロッパ最古層(クロマニョン人)の集団にもそのハプログループに属す者がいた。もう少し遡ると、インドやアラビアなどに散見されるC1b1(M356)、インドネシア東部からメラネシアおよびポリネシア、オーストラリアの先住民アボリジニに多く見られるC1b3(B477)との共通祖先C1にたどり着く。
崎谷満はC1a1の祖型はイラン付近からアルタイ山脈付近を経由し朝鮮半島経由で日本に到達したとしている[8]。https://ja.wikipedia.org/wiki/%E3%83%8F%E3%83%97%E3%83%AD%E3%82%B0%E3%83%AB%E3%83%BC%E3%83%97M7a_(mtDNA)
古人骨
鳥取県鳥取市青谷上寺地遺跡で発掘された弥生時代後期とみられる人骨について、県が国立科学博物館や国立歴史民俗博物館が分析した父系結果、4点のうち2点はハプログループC1a1(M8)であった[9]。
ーーーーーーー
C1aの人々はC1bの人々とともにスンダランドに移り住んでいた。氷河時代とは言え暖かく、食料には苦労しなかったであろう。しかし、最終氷期が終わり、海水面が上昇し、次第にスンダランドが水没してきた。人々は船に乗って移動することとなった。C1aの人々は黒潮に乗って北上し、沖縄にたどり着いた。17000年ほど前のことである。港川人である。 この頃は黒潮が九州の南側を流れており、当時の船では九州にたどり着くことができなかった。
しかし、15000年ほど前に対馬海峡が広がり、黒潮から対馬海流が分流するようになり、舟で南九州に流れ着くことができるようになり、南九州にたどり着いた。C1a1系統の人物は日本列島固有であり、現代日本人に5%ほど存在している。C1a系統は40000年ほど前に発現しているが、その子系統の拡散が始まったのが12000年ほど前からであり、その間に長いギャップがある。これより、日本列島流入時期が不確定となるが、拡散が始まる少し前に流入したと考えている。C1a1系統に近いC1a2がヨーロッパやアルメニアに多い。その間の地域に存在せず、その経路が特定できない。近い系統のC1bの多いスンダランドから来たと推定しているが、人数も少なく、スンダランドに残ったC1a1系統は絶滅したのではないだろうか。
港川人のハブログループは確認されていないが、出土した人骨の骨格がオーストラリアの原住民と似ているので、スンダランドから来た系統と思われ、C1a1系統と推定する。
ーーーーーーー
…日本人の遺伝子の半分は、ユーラシア大陸で最古の遺伝子(C1a1・D1b)であり、
残りの半分は最も新しい種類の遺伝子(O1b2・O2)なのです。
このようなバランスを持つ地域は、地球上でも本当に稀です。
そのせいか、この国には、最も古いものが、最も新しいものと無理なく自然に共存するという、世界に類のない特性が、至る所で見られます。融和と共生の精神が息づいています。世界の絶滅危惧種であるD系統ハプログループが、この国にこれほど多く生き残っていることの意味を、深く考えてみてください。
この国に深く根付いているポリシーは、「決して敵を滅ぼさない」「根絶やしにしない」ということです。「敵を滅ぼす者は、自らも滅ぼされる。」わたしたちの祖先は、その真理を歴史からも学び、その知恵を「平家物語」「太平記」のようなカタチでも語り継いできました。
ーーーーーーー
7300年前~5500年前
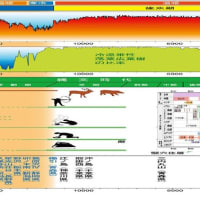
縄文前期から中期にかけては最も典型的な縄文文化が栄えた時期であり、現在は三内丸山遺跡と呼ばれる場所に起居した縄文人たちが保持していたのも、主にこの時期の文化形式である。海水面は縄文前期の中頃には現在より3mほど高くなり、気候も現在より温暖であった。この時期のいわゆる縄文海進によって沿岸部には好漁場が増え、海産物の入手も容易になった。しかし、縄文早期から前期に代わるときに大異変が起こっている。鬼界カルデラ噴火である。
南九州先進文化
鹿児島県上野原遺跡で焼石による蒸焼きや燻製のを目的とする集石遺構や連結坑が見つかった。これらは南方系の文化であり、他に、貝殻文土器、壺、耳飾り、石斧などが多数見つかっている。貝殻文土器は縄の代わりに貝殻で文様を付けた土器である。壺は、本州では縄文後期から晩期にならないと現われない。土器のリング状の耳飾りも本州より4000年も早い。石斧は丸木舟を作るためのもので、フィリピンや台湾、沖縄で似たものが見つかっている。
この頃南九州に住んでいたのは、スンダランド出身で南方系のミトコンドリアハブログループM7系統で、南西諸島を伝って、18000年ほど前に上陸した人々であろう。Y染色体ハプログループではC1a1系統と思われる。
およそ1万年前から始まった南九州の縄文早期文化は、当時の日本列島では最先端であったことがわかる。ところが、7300年前の鬼界カルデラ噴火によって、この最先端文化は壊滅したのである。
南九州の先進性を示したのが図7である。列島の縄文文化に先駆けた南九州の縄文文化は先に述べたように7300年前の鬼界カルデラのアカホヤ噴火で壊滅した。そこに住んでいた縄文人は生き延びて他の地域に移り住んだようである。それは、図7の左上の文化が2000~4000年を経て右下に伝えられていることからも解る。
鬼界カルデラ噴火
7300年前、鬼界カルデラの超巨大噴火が起きた。鬼界カルデラは屋久島の北にある。 25×15 ㎞の大きさで、薩摩硫黄島や竹島が外輪山である。この噴火は日本列島で最も新しい超巨大噴火である。噴出量170 ㎞3以上でアカホヤ噴火ともよばれる。火砕流は海上を走り、屋久島、種子島、九州南部を覆った。火山灰は東北南部まで達し、紀伊や山陽で20㎝を超えたと推定されている。宮崎県南部では60㎝を超えたとされている。
この火砕流で、南九州に住んでいた縄文人は壊滅したようである。そこに住んでいた縄文人の一部は生き延びて他の地域に移り住んだようである。それは、南九州の先進文化が2000~4000年を経て本州に伝えられていることからも解る。そして、以後1000年間人が住んだ形跡が見られない。その後に住み着いた前期縄文人の系統(D1a2a)は今までの系統(C1a1)と異なるようである。
ーーーーーーー
Y染色体C1系統(縄文系)
沖縄県具志頭村から出土した古代人骨・港川人(約1万7000年前)は、オーストラリア先住民と骨格が近似しているため、Y染色体はハプログループC系統のC1a1(C-M8)に連なる系統にあたると推定。
Y染色C1系統は、YAP因子を持たない縄文人です。