








【眼の誕生1】
友人のW君からこういうメールが来た。
<目の発生に疑問をもっていたのですが、NHK特集でロドプシン遺伝子はミドリムシが最初としってびっくりしました。昆虫の複眼と魚類のカメラ眼とどこでわかれたかご存知ですか?>
私は問題の番組を見ていないが、すこしこの問題を考えてみたい。
1)走光性(フォトタキシス)の起源=
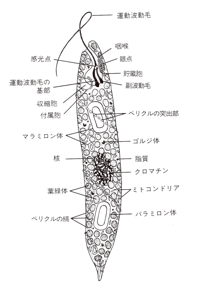
図は「細胞内小器官の寄生体由来説」を唱えた生物学者リン・マーグリス『五つの王国』(日経サイエンス社, 1987)にあるミドリムシの模式図だ(図1)。
(図1)
(図2:拡大図)

ミドリムシは葉緑体、ミトコンドリア、核、鞭毛という細胞内小器官があり、真核生物で光合成をしながら、同時にアメーバのように食物を取り込むことができるという、便利な単細胞だ。図1の上方が頭(前端)で、ここに「運動波動毛(鞭毛)」がある。この主毛をらせん状に動かして前に進む。
この鞭毛は「咽喉」と呼ばれる袋状の構造物の奥壁から出ている。ミドリムシは「光走性(phototaxis)」といって、太陽光のある方向に動く。これは光合成をするためだ。しかし、主毛だけでは向きを調整できないので、「副波動毛」というもう1本の鞭毛が、主毛の根元に付着していて、これが主毛の動きを調節して、動く方向をコントロールしている。
咽喉の副波動毛側に、「眼点(eyespot)」があり、これはカロテノイドを含んだ顆粒の集合体である。このため眼点は赤橙色を呈し、長い間「眼」だと誤認されてきたが、実際には鞭毛が副波動毛に接触する位置にある、「傍鞭毛体(para-flagellar body)」が「感光点(photosensitive spot)」である。
最近になって解明されたことなので、「岩波生物学辞典・第5版」(2013)の記載は正確だが、WIKI(英語、日本語)とも記載が混乱している。
つまりいわゆる「眼点」は実際には感光点を円蓋状に被い、光を吸収することで感光点に当たる光が、ベクトルを持つように作用している。
この感光点での光化学反応については、院内感染で問題となる緑膿菌が代表種である、緑藻類シュウドモナス(Pseudomonas)目の場合は、傍鞭毛体にある感光素はオプシンタンパクにVit. A(レチナール)が結合した「ロドプシン」だとされている。
他方で、ユーグレナ目ミドリムシの感光素は、青色により活性化するアデニール・サイクレースだという。(英語WIKIのM.Iseki, 2002論文)この点はNHKが報じたとされる、「古代型ロドプシン」とは異なっており、確認が必要だろう。
光を検知するには、光子(フォトン)のエネルギーを吸収して立体異性体に変わる発色団が必要だ。発色団が異なっていても、その後の化学反応には共通性があり、活性型の中間体を経て、「Gタンパク質」が活性化され、そこでcAMPかcGMPが「細胞内信号伝達」物質として形成される。このシグナル物質が細胞膜を介してのNa/Kイオン移動に影響を与え、光が電気信号に変わるという点では共通している。
この光化学反応が介在するおかげで、ロドプシン1分子にエネルギーを与えた光子(フォトン)1個の作用は、50万倍に増幅されるという(後藤祐児ほか『タンパク質化学』化学同人,2006)
但し、疑問点があって、感光点で光が電気信号に変換されるとすれば、それがどうやって主鞭毛を動かす、副鞭毛の基底小体にある微小細管という収縮性タンパクに伝えられるのか、副鞭毛の動きがどのように主鞭毛の動きを調節するのかが、私にはよくわからない。
2)眼点が眼の原基である=
最初の真核細胞が出現したのは12億年前とされている。核をもたない原核細胞の大腸菌にすでに現存する遺伝子の多くが認められることを考えると、クラミドモナスやミドリムシに認められる「光=電気信号」変換装置は、おそらく「最初の眼」と言ってよいのではないか。
最初に「眼」が登場したのは、扁形動物のプラナリアにおいてで、多細胞動物でも二胚葉性の海綿動物やクラゲなどの刺胞動物では、単細胞性の明暗識別細胞に留まっていたようだ。ともかく、6億5000万年前から8000万年続いた「エディアカラ期」(オーストラリアにある地層の名前に由来)に初めて左右対称性の眼をもつプラナリアが出現した。
分子生物学から進化の問題に取り組んだ大野乾は「進化の原動力は遺伝子重複にある」と唱えた(『生命の誕生と進化』,東大出版会, 1988)。
単細胞のミドリムシに認められた、「凹んだ袋の外側に光をさえぎる色素層があり、袋の内側に光感受性の細胞末端がある」という眼点の構造は、プラナリアでもそっくりそのままに、再現されている。但し多細胞レベルで…。(図3:岩堀修明『感覚器の進化』,ブルーバックス, 2011)
これは「遺伝子重複」の結果としか考えられない。
 (図3)
(図3)
面白いことに、この眼は凹状(窩状)をしていて、壁にはメラニンを持つ色素細胞の層がある。視神経は前から進入し、色素細胞層の前で「光受容部」にあたる細胞突起を扇状に拡げている。
論理的には色素細胞層が哺乳類の網膜の色素細胞層に相当する。視神経が色素細胞層の反対側から進入して、凹部壁内面に拡がれば「網膜」になる。眼の前側にレンズをつければ、哺乳類の眼ができる。
つまりミドリムシの眼点とプラナリアの眼とヒトの眼は「相同」というわけだ。
プラナリアの「眼」は、光の進入方向と視神経の進入方向が同じで、光受容部が固定されていないから、光刺激の度合いと光源の方向は高感度にとらえることができるが、光を「像」として把握することはできない。
色素細胞層と視神経の関係が逆転して、色素細胞の後から視神経が進入するようになったものがアンモナイトやアワビ(軟体動物頭足類)の「窩状眼」である。この原理はレンズのないピンホールカメラにフィルムを取り付けたものと同じである。真の網膜はここで初めて出現した。
このタイプの「単眼」は4億6500万年前から5000万年続いた「デヴォン紀」(英国南部海岸の地名に由来)に初めて出現した。
同じ頭足類でもイカやタコでは、角膜、水晶体と毛様体を備えた「水晶体眼」が出現する。
軟体動物の水晶体眼は、目蓋まで備えていて、構造上は脊椎動物の水晶体眼と同じである。(厳密には色素細胞層の位置が異なる)
ハーバードの進化生物学者S.J.グールドはこれを「進化の収斂」として説明したが、両者の類似は「収斂」ではなく、5億7000万年前に始まり7000万年続いた「カンブリア大爆発」における生物多様化の結果と見なすのが妥当であろう。(カンブリアは英国ウェールズの古称に由来する地質年代名)。つまり水晶眼を形成するのに必要な遺伝子はこの時期に創出され、最初、軟体動物頭足類で発現され、後に脊索動物で発現されたにすぎないであろう。
3)複眼の進化=
複眼の出発点は二胚葉動物のヒドラに認められる。外上皮細胞内に散在する感覚細胞である。(図4: Buchsbaumから)ヒドラでは上皮細胞は「上皮=筋細胞」と呼ばれ、内側では細胞下部に筋繊維が輪状となり、外側のものは縦に走っている。(ヒト腸管に認められる筋繊維の配置「内輪縦走」のプロトタイプはここにすでに存在する。)
この外層上皮細胞の間に介在しているのが、Sensory cell(感覚細胞)で、基底部から突起を出し神経細胞とつながり、原始的な神経系を形成している。この感覚細胞にロドプシン遺伝子が発現し、Gタンパク質—cAMPないしcGMPシステムにより光信号を電気信号に変える。
 (図4)
(図4)
この感覚細胞の外側に透明化がおき、水晶体(ないし硝子体)が形成され、上部に光感覚部が移動し、細胞質と核が下方に移動し、基底部から神経線維(電気信号伝達装置)が出るような配置転換をしたものが、もっとも単純な「眼」(「個眼」と呼ばれる)である。
この視覚細胞がユニットとして量的に複合したものが、初期の複眼である。その場合、一個のユニットから光の漏洩を防ぐために、円柱状のユニットを取りまいて遮光する色素(メラニン)細胞によるラッピングが必要となる。
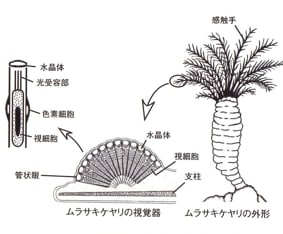
この原始的な複眼は基本的には定着性の環形動物、溝副触手綱のムラサキケヤリに認められる。(図5:岩堀から)ムラサキケヤリは泥で造った住居に胴体を入れ、触手を海水中に突き出している。
 (図5)
(図5)
個々の触手に多数の「複眼」があるが、それは図5左上のような単純な「管状眼」が集合したものである。
扁形動物のプラナリアの眼では、光の進入方向と視細胞から電気信号が出る方向は逆向きであったが、環形動物のムラサキケヤリの単眼では光の進入方向に向かって視細胞が配置されている。このような「眼の軸」の逆転は、細胞配置を決めるより上位の「調節遺伝子」が出現したために起こったものであろう。
複眼を構成する「個眼」の構造も、エディアカラ紀に環形動物多毛類(ゴカイなど)に最初に出現したものは、単純な管状眼だが、それに続いたカンブリア紀に出現した節足動物(昆虫、甲殻類、三葉虫など)では、個眼そのものが角膜、水晶体、複数の視細胞からなり、このユニットが遠位、近位にある2種のメラニン細胞によりラップされたものからなる、一種の器官(完全複眼)としてよく発達したようだ。
この複雑な個眼(水晶体眼で視神経が脳の視覚中枢に繋がっていない点を除けば、脊椎動物の眼に匹敵する)が、少なくとも10個(アリ)、多いもの(ハエ、トンボ)では1~2.8万個集合して「複眼」を形成している。これは小さいサイズの個眼が多数集まったものなので「集合複眼」と呼ばれる。
英国の進化生物学者A.パーカーは、三葉虫の最古の化石にすでに完全複眼が認められことから、「結像能力」を持つ最初の眼は、5億4300万年前、カンブリア初期に出現した最初の三葉虫が備えていた、一対の完全複眼だと主張している。(『眼の誕生』, 草思社, 2006)
このタイプの単眼に、角膜が付き、レンズが付き、虹彩が付くと、ヒトを含めた哺乳類の眼球が出現する。タコの水晶眼から脊椎動物の眼が進化したとすれば、物語は単純なのだが、じつはそうでない。
4)原索—脊椎動物の眼=
さてW君の質問「昆虫の複眼と魚類のカメラ眼とどこでわかれたかご存知ですか?」に回答するには、昆虫は無脊椎動物であり、脊椎動物・魚類の出発点は、ホヤやナメクジウオなどが属する原索動物であることに、触れないわけにいかない。(以下は次回「眼の誕生2」にゆずる。)
友人のW君からこういうメールが来た。
<目の発生に疑問をもっていたのですが、NHK特集でロドプシン遺伝子はミドリムシが最初としってびっくりしました。昆虫の複眼と魚類のカメラ眼とどこでわかれたかご存知ですか?>
私は問題の番組を見ていないが、すこしこの問題を考えてみたい。
1)走光性(フォトタキシス)の起源=
図は「細胞内小器官の寄生体由来説」を唱えた生物学者リン・マーグリス『五つの王国』(日経サイエンス社, 1987)にあるミドリムシの模式図だ(図1)。
(図1)
(図2:拡大図)
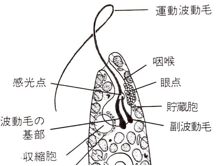
ミドリムシは葉緑体、ミトコンドリア、核、鞭毛という細胞内小器官があり、真核生物で光合成をしながら、同時にアメーバのように食物を取り込むことができるという、便利な単細胞だ。図1の上方が頭(前端)で、ここに「運動波動毛(鞭毛)」がある。この主毛をらせん状に動かして前に進む。
この鞭毛は「咽喉」と呼ばれる袋状の構造物の奥壁から出ている。ミドリムシは「光走性(phototaxis)」といって、太陽光のある方向に動く。これは光合成をするためだ。しかし、主毛だけでは向きを調整できないので、「副波動毛」というもう1本の鞭毛が、主毛の根元に付着していて、これが主毛の動きを調節して、動く方向をコントロールしている。
咽喉の副波動毛側に、「眼点(eyespot)」があり、これはカロテノイドを含んだ顆粒の集合体である。このため眼点は赤橙色を呈し、長い間「眼」だと誤認されてきたが、実際には鞭毛が副波動毛に接触する位置にある、「傍鞭毛体(para-flagellar body)」が「感光点(photosensitive spot)」である。
最近になって解明されたことなので、「岩波生物学辞典・第5版」(2013)の記載は正確だが、WIKI(英語、日本語)とも記載が混乱している。
つまりいわゆる「眼点」は実際には感光点を円蓋状に被い、光を吸収することで感光点に当たる光が、ベクトルを持つように作用している。
この感光点での光化学反応については、院内感染で問題となる緑膿菌が代表種である、緑藻類シュウドモナス(Pseudomonas)目の場合は、傍鞭毛体にある感光素はオプシンタンパクにVit. A(レチナール)が結合した「ロドプシン」だとされている。
他方で、ユーグレナ目ミドリムシの感光素は、青色により活性化するアデニール・サイクレースだという。(英語WIKIのM.Iseki, 2002論文)この点はNHKが報じたとされる、「古代型ロドプシン」とは異なっており、確認が必要だろう。
光を検知するには、光子(フォトン)のエネルギーを吸収して立体異性体に変わる発色団が必要だ。発色団が異なっていても、その後の化学反応には共通性があり、活性型の中間体を経て、「Gタンパク質」が活性化され、そこでcAMPかcGMPが「細胞内信号伝達」物質として形成される。このシグナル物質が細胞膜を介してのNa/Kイオン移動に影響を与え、光が電気信号に変わるという点では共通している。
この光化学反応が介在するおかげで、ロドプシン1分子にエネルギーを与えた光子(フォトン)1個の作用は、50万倍に増幅されるという(後藤祐児ほか『タンパク質化学』化学同人,2006)
但し、疑問点があって、感光点で光が電気信号に変換されるとすれば、それがどうやって主鞭毛を動かす、副鞭毛の基底小体にある微小細管という収縮性タンパクに伝えられるのか、副鞭毛の動きがどのように主鞭毛の動きを調節するのかが、私にはよくわからない。
2)眼点が眼の原基である=
最初の真核細胞が出現したのは12億年前とされている。核をもたない原核細胞の大腸菌にすでに現存する遺伝子の多くが認められることを考えると、クラミドモナスやミドリムシに認められる「光=電気信号」変換装置は、おそらく「最初の眼」と言ってよいのではないか。
最初に「眼」が登場したのは、扁形動物のプラナリアにおいてで、多細胞動物でも二胚葉性の海綿動物やクラゲなどの刺胞動物では、単細胞性の明暗識別細胞に留まっていたようだ。ともかく、6億5000万年前から8000万年続いた「エディアカラ期」(オーストラリアにある地層の名前に由来)に初めて左右対称性の眼をもつプラナリアが出現した。
分子生物学から進化の問題に取り組んだ大野乾は「進化の原動力は遺伝子重複にある」と唱えた(『生命の誕生と進化』,東大出版会, 1988)。
単細胞のミドリムシに認められた、「凹んだ袋の外側に光をさえぎる色素層があり、袋の内側に光感受性の細胞末端がある」という眼点の構造は、プラナリアでもそっくりそのままに、再現されている。但し多細胞レベルで…。(図3:岩堀修明『感覚器の進化』,ブルーバックス, 2011)
これは「遺伝子重複」の結果としか考えられない。
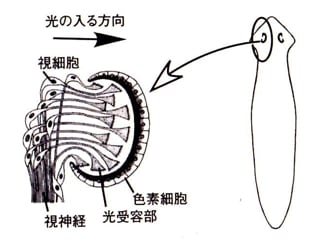 (図3)
(図3) 面白いことに、この眼は凹状(窩状)をしていて、壁にはメラニンを持つ色素細胞の層がある。視神経は前から進入し、色素細胞層の前で「光受容部」にあたる細胞突起を扇状に拡げている。
論理的には色素細胞層が哺乳類の網膜の色素細胞層に相当する。視神経が色素細胞層の反対側から進入して、凹部壁内面に拡がれば「網膜」になる。眼の前側にレンズをつければ、哺乳類の眼ができる。
つまりミドリムシの眼点とプラナリアの眼とヒトの眼は「相同」というわけだ。
プラナリアの「眼」は、光の進入方向と視神経の進入方向が同じで、光受容部が固定されていないから、光刺激の度合いと光源の方向は高感度にとらえることができるが、光を「像」として把握することはできない。
色素細胞層と視神経の関係が逆転して、色素細胞の後から視神経が進入するようになったものがアンモナイトやアワビ(軟体動物頭足類)の「窩状眼」である。この原理はレンズのないピンホールカメラにフィルムを取り付けたものと同じである。真の網膜はここで初めて出現した。
このタイプの「単眼」は4億6500万年前から5000万年続いた「デヴォン紀」(英国南部海岸の地名に由来)に初めて出現した。
同じ頭足類でもイカやタコでは、角膜、水晶体と毛様体を備えた「水晶体眼」が出現する。
軟体動物の水晶体眼は、目蓋まで備えていて、構造上は脊椎動物の水晶体眼と同じである。(厳密には色素細胞層の位置が異なる)
ハーバードの進化生物学者S.J.グールドはこれを「進化の収斂」として説明したが、両者の類似は「収斂」ではなく、5億7000万年前に始まり7000万年続いた「カンブリア大爆発」における生物多様化の結果と見なすのが妥当であろう。(カンブリアは英国ウェールズの古称に由来する地質年代名)。つまり水晶眼を形成するのに必要な遺伝子はこの時期に創出され、最初、軟体動物頭足類で発現され、後に脊索動物で発現されたにすぎないであろう。
3)複眼の進化=
複眼の出発点は二胚葉動物のヒドラに認められる。外上皮細胞内に散在する感覚細胞である。(図4: Buchsbaumから)ヒドラでは上皮細胞は「上皮=筋細胞」と呼ばれ、内側では細胞下部に筋繊維が輪状となり、外側のものは縦に走っている。(ヒト腸管に認められる筋繊維の配置「内輪縦走」のプロトタイプはここにすでに存在する。)
この外層上皮細胞の間に介在しているのが、Sensory cell(感覚細胞)で、基底部から突起を出し神経細胞とつながり、原始的な神経系を形成している。この感覚細胞にロドプシン遺伝子が発現し、Gタンパク質—cAMPないしcGMPシステムにより光信号を電気信号に変える。
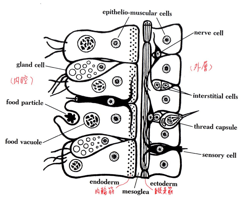 (図4)
(図4) この感覚細胞の外側に透明化がおき、水晶体(ないし硝子体)が形成され、上部に光感覚部が移動し、細胞質と核が下方に移動し、基底部から神経線維(電気信号伝達装置)が出るような配置転換をしたものが、もっとも単純な「眼」(「個眼」と呼ばれる)である。
この視覚細胞がユニットとして量的に複合したものが、初期の複眼である。その場合、一個のユニットから光の漏洩を防ぐために、円柱状のユニットを取りまいて遮光する色素(メラニン)細胞によるラッピングが必要となる。
この原始的な複眼は基本的には定着性の環形動物、溝副触手綱のムラサキケヤリに認められる。(図5:岩堀から)ムラサキケヤリは泥で造った住居に胴体を入れ、触手を海水中に突き出している。
 (図5)
(図5) 個々の触手に多数の「複眼」があるが、それは図5左上のような単純な「管状眼」が集合したものである。
扁形動物のプラナリアの眼では、光の進入方向と視細胞から電気信号が出る方向は逆向きであったが、環形動物のムラサキケヤリの単眼では光の進入方向に向かって視細胞が配置されている。このような「眼の軸」の逆転は、細胞配置を決めるより上位の「調節遺伝子」が出現したために起こったものであろう。
複眼を構成する「個眼」の構造も、エディアカラ紀に環形動物多毛類(ゴカイなど)に最初に出現したものは、単純な管状眼だが、それに続いたカンブリア紀に出現した節足動物(昆虫、甲殻類、三葉虫など)では、個眼そのものが角膜、水晶体、複数の視細胞からなり、このユニットが遠位、近位にある2種のメラニン細胞によりラップされたものからなる、一種の器官(完全複眼)としてよく発達したようだ。
この複雑な個眼(水晶体眼で視神経が脳の視覚中枢に繋がっていない点を除けば、脊椎動物の眼に匹敵する)が、少なくとも10個(アリ)、多いもの(ハエ、トンボ)では1~2.8万個集合して「複眼」を形成している。これは小さいサイズの個眼が多数集まったものなので「集合複眼」と呼ばれる。
英国の進化生物学者A.パーカーは、三葉虫の最古の化石にすでに完全複眼が認められことから、「結像能力」を持つ最初の眼は、5億4300万年前、カンブリア初期に出現した最初の三葉虫が備えていた、一対の完全複眼だと主張している。(『眼の誕生』, 草思社, 2006)
このタイプの単眼に、角膜が付き、レンズが付き、虹彩が付くと、ヒトを含めた哺乳類の眼球が出現する。タコの水晶眼から脊椎動物の眼が進化したとすれば、物語は単純なのだが、じつはそうでない。
4)原索—脊椎動物の眼=
さてW君の質問「昆虫の複眼と魚類のカメラ眼とどこでわかれたかご存知ですか?」に回答するには、昆虫は無脊椎動物であり、脊椎動物・魚類の出発点は、ホヤやナメクジウオなどが属する原索動物であることに、触れないわけにいかない。(以下は次回「眼の誕生2」にゆずる。)












※コメント投稿者のブログIDはブログ作成者のみに通知されます