








栽培植物の起源と伝播 No9
遺伝学の手法を使ったドエブリーがトウモロコシの祖先を絞り込む
メキシコ、グアテマラ、ニカラグアに生息する5つの野生種テオシントがトウモロコシの起源に関係したことが明らかになり、その中ではバルサバレー(Balsas River Valley)で発見された野生種のバルサ・テオシント(学名:Zea mays subsp. parviglumis)が最もトウモロコシに近いということがわかった。
この野生種のバルサ・テオシントを発見したのは、ウイスコンシン・マディソン大学の植物学教授イルチスとこの大学で博士課程を受けていたドエブリーで、1977年9月22日のことだった。
1980年に博士課程を終了したドエブリーは、ノースカロライナ州立大学に移り、遺伝学者メジャー・グッドマン(Major Goodman)の指導の下で野生種のテオシントとトウモロコシの進化の関係を調査・研究した。どんな方法で調べたかは後で説明することにして、誰もが想像もしなかった結果を発見した。
それは、 『バルサ・テオシント(学名:Zea mays subsp. parviglumis)が遺伝的にトウモロコ(Zea mays subsp. mays)と最も近い。』 ということだった。
言い換えると、現代のトウモロコシの祖先は、バルサ川流域を原産地とする野生のテオシント、パヴィルミス(学名がZea mays subsp. Parviglumis)であり、古代の遊牧民がこの種を栽培し始めて今日の洗練したトウモロコシに至った。ということになる。
見た目の形態が大きく異なるパヴィルミスとトウモロコシなので“信じられない”というのが当然で、ドエブリーの遺伝子学からの主張が正しいのならば、「古代の遊牧民がパヴィルミスを栽培した」という痕跡がどこかにあるはずだ。
ドエブリーが主張した時期に考古学的にわかっていたことは、メキシコシティの南東部にあるテワカンバレーにあるコスカトラン洞窟(Coxcatlan Cave)で8千年前(紀元前5960年頃)に栽培された植物(トウモロコシ、ヒョウタン、スカッシュ、豆)の痕跡を見つけ、さらにテワカンバレーから160km南東に行ったところにオアハカバレーにあるギラ・ナキツ(Guilá Naquitz)洞窟で、テワカンバレーで発見したトウモロコシよりも700年も古いトウモロコシを発掘した。
このトウモロコシは、メキシカーナ(学名:Zea mays subsp. Mexicana)のようであり、この種が現在のトウモロコシの祖先に近い種とみなされていた。
パヴィルミスとメキシカーナ2種の気候的・地理的な位置関係を表したものが下記の地図になる。
メキシカーナは、寒冷な熱帯高地で生育しテワカンバレーはこの気候帯にあり、パヴィルミスは暑い熱帯低地で生育しバルサスバレーはここにある。
1970年代までは、化石などの保存状態が良い亜熱帯の乾燥した地域か寒冷な熱帯高地での考古学発掘しかされていなかったので熱帯低地は手がついていなかった。
それにしても、この2種は近いところで生育し交雑した可能性もある。
(地図)パヴィルミスとメキシカーナの地理的分布
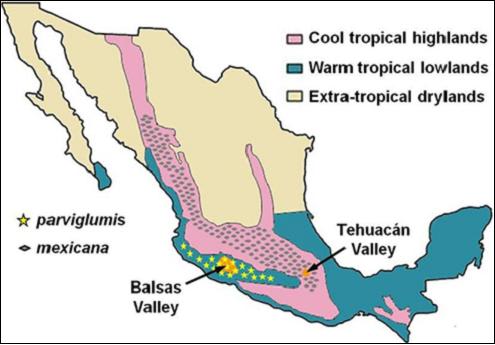
(出典) McClung de Tapia (1992) and Matsuoka et al. (2002).
遺伝子の簡単な歴史
遺伝子の概念は、現在のチェコのブルノの司祭だったメンデル(Gregor Johann Mendel、1822-1884)によって1865年に発見された。えんどう豆の交配を繰り返し、背の高さなどの表現形質が異なるものに注目し、必ず背が高く育つ種子と、必ず背が低く育つ種子を選別し、これらを交配させてみると必ず背が高く育った。これを優性の法則というが、親から子に伝わる遺伝的な粒子があることを確信した。
後にこの粒子をウィリアム・ベイトソン(William Bateson, 1861-1926)によって1905年に遺伝子と名づけられたが、それまではこのメンデルの法則は忘れられていた。
遺伝子学が大きく進んだのは、1953年のジェームス・ワトソン(James Dewey Watson, 1928- )とフランシス・クリック(Francis Harry Compton Crick, 1916-2004)によってDNAの二重らせん構造モデルが発表されてからで、1966年にはDNAの暗号解読が完了した。
ドエブリーの手法
ノースカロライナ州立大学でのドエブリーの研究は、野生種のテオシントがトウモロコシに進化していく過程を調べるために異系酵素(allozymes)を使って調査した。
この異系酵素(一種の遺伝子マーカー)は、進化の履歴と生物の異なる種の関係を測定する時に使われる生物学的な酵素で、この酵素のアミノ酸配列を比較することによって親子などの関係がわかるという。
一種の遺伝子マーカーとして酵素のアミノ酸配列を使用したというが、遺伝子マーカーは一般的にはDNA型鑑定として知られている。
いまではサスペンスドラマを見ると犯罪捜査の必需品であり、タバコの吸い口についた唾液、血痕、髪の毛などからDNAの配列を調べ、犯人が残した遺留品に付着した血痕などのDNAと比較して同一か否かを完璧ではないが100%に近い確率で判定できる。
また親子の関係を調べるのにも使われていて、生物個体の遺伝的性質と系統(個人の特定、親子関係、親族関係、祖先など)の目印になるという。
(写真)(a)トウモロコシ(Zea mays subsp. mays)(b)パヴィルミス(Zea mays subsp. parviglumis)

(出典) ゲノムの変化からみた トウモロコシの栽培化と育種by山崎将紀
ドエブリー以前の種の同定は、雄しべ・雌しべ、花びらの数・形、茎・葉などの生え方など外観でわかる形態などによっていたので、野生種のパヴィルミスはとても現在のトウモロコシの祖先とは考えることすら出来なかった。
パヴィルミス(Zea mays subsp. Parviglumis)がトウモロコシの祖先だとする遺伝子学からの結論が正しいのならば、古代の遊牧民により栽培され食された痕跡がどこかにあるはずで、この発見と年代の測定により証明することが出来ることになる。
遺伝学の手法を使ったドエブリーがトウモロコシの祖先を絞り込む
メキシコ、グアテマラ、ニカラグアに生息する5つの野生種テオシントがトウモロコシの起源に関係したことが明らかになり、その中ではバルサバレー(Balsas River Valley)で発見された野生種のバルサ・テオシント(学名:Zea mays subsp. parviglumis)が最もトウモロコシに近いということがわかった。
この野生種のバルサ・テオシントを発見したのは、ウイスコンシン・マディソン大学の植物学教授イルチスとこの大学で博士課程を受けていたドエブリーで、1977年9月22日のことだった。
1980年に博士課程を終了したドエブリーは、ノースカロライナ州立大学に移り、遺伝学者メジャー・グッドマン(Major Goodman)の指導の下で野生種のテオシントとトウモロコシの進化の関係を調査・研究した。どんな方法で調べたかは後で説明することにして、誰もが想像もしなかった結果を発見した。
それは、 『バルサ・テオシント(学名:Zea mays subsp. parviglumis)が遺伝的にトウモロコ(Zea mays subsp. mays)と最も近い。』 ということだった。
言い換えると、現代のトウモロコシの祖先は、バルサ川流域を原産地とする野生のテオシント、パヴィルミス(学名がZea mays subsp. Parviglumis)であり、古代の遊牧民がこの種を栽培し始めて今日の洗練したトウモロコシに至った。ということになる。
見た目の形態が大きく異なるパヴィルミスとトウモロコシなので“信じられない”というのが当然で、ドエブリーの遺伝子学からの主張が正しいのならば、「古代の遊牧民がパヴィルミスを栽培した」という痕跡がどこかにあるはずだ。
ドエブリーが主張した時期に考古学的にわかっていたことは、メキシコシティの南東部にあるテワカンバレーにあるコスカトラン洞窟(Coxcatlan Cave)で8千年前(紀元前5960年頃)に栽培された植物(トウモロコシ、ヒョウタン、スカッシュ、豆)の痕跡を見つけ、さらにテワカンバレーから160km南東に行ったところにオアハカバレーにあるギラ・ナキツ(Guilá Naquitz)洞窟で、テワカンバレーで発見したトウモロコシよりも700年も古いトウモロコシを発掘した。
このトウモロコシは、メキシカーナ(学名:Zea mays subsp. Mexicana)のようであり、この種が現在のトウモロコシの祖先に近い種とみなされていた。
パヴィルミスとメキシカーナ2種の気候的・地理的な位置関係を表したものが下記の地図になる。
メキシカーナは、寒冷な熱帯高地で生育しテワカンバレーはこの気候帯にあり、パヴィルミスは暑い熱帯低地で生育しバルサスバレーはここにある。
1970年代までは、化石などの保存状態が良い亜熱帯の乾燥した地域か寒冷な熱帯高地での考古学発掘しかされていなかったので熱帯低地は手がついていなかった。
それにしても、この2種は近いところで生育し交雑した可能性もある。
(地図)パヴィルミスとメキシカーナの地理的分布
(出典) McClung de Tapia (1992) and Matsuoka et al. (2002).
遺伝子の簡単な歴史
遺伝子の概念は、現在のチェコのブルノの司祭だったメンデル(Gregor Johann Mendel、1822-1884)によって1865年に発見された。えんどう豆の交配を繰り返し、背の高さなどの表現形質が異なるものに注目し、必ず背が高く育つ種子と、必ず背が低く育つ種子を選別し、これらを交配させてみると必ず背が高く育った。これを優性の法則というが、親から子に伝わる遺伝的な粒子があることを確信した。
後にこの粒子をウィリアム・ベイトソン(William Bateson, 1861-1926)によって1905年に遺伝子と名づけられたが、それまではこのメンデルの法則は忘れられていた。
遺伝子学が大きく進んだのは、1953年のジェームス・ワトソン(James Dewey Watson, 1928- )とフランシス・クリック(Francis Harry Compton Crick, 1916-2004)によってDNAの二重らせん構造モデルが発表されてからで、1966年にはDNAの暗号解読が完了した。
ドエブリーの手法
ノースカロライナ州立大学でのドエブリーの研究は、野生種のテオシントがトウモロコシに進化していく過程を調べるために異系酵素(allozymes)を使って調査した。
この異系酵素(一種の遺伝子マーカー)は、進化の履歴と生物の異なる種の関係を測定する時に使われる生物学的な酵素で、この酵素のアミノ酸配列を比較することによって親子などの関係がわかるという。
一種の遺伝子マーカーとして酵素のアミノ酸配列を使用したというが、遺伝子マーカーは一般的にはDNA型鑑定として知られている。
いまではサスペンスドラマを見ると犯罪捜査の必需品であり、タバコの吸い口についた唾液、血痕、髪の毛などからDNAの配列を調べ、犯人が残した遺留品に付着した血痕などのDNAと比較して同一か否かを完璧ではないが100%に近い確率で判定できる。
また親子の関係を調べるのにも使われていて、生物個体の遺伝的性質と系統(個人の特定、親子関係、親族関係、祖先など)の目印になるという。
(写真)(a)トウモロコシ(Zea mays subsp. mays)(b)パヴィルミス(Zea mays subsp. parviglumis)
(出典) ゲノムの変化からみた トウモロコシの栽培化と育種by山崎将紀
ドエブリー以前の種の同定は、雄しべ・雌しべ、花びらの数・形、茎・葉などの生え方など外観でわかる形態などによっていたので、野生種のパヴィルミスはとても現在のトウモロコシの祖先とは考えることすら出来なかった。
パヴィルミス(Zea mays subsp. Parviglumis)がトウモロコシの祖先だとする遺伝子学からの結論が正しいのならば、古代の遊牧民により栽培され食された痕跡がどこかにあるはずで、この発見と年代の測定により証明することが出来ることになる。







※コメント投稿者のブログIDはブログ作成者のみに通知されます