獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2025年3月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | ||||||||
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| 9 | 10 | 11 | 12 | 13 | 14 | 15 | ||
| 16 | 17 | 18 | 19 | 20 | 21 | 22 | ||
| 23 | 24 | 25 | 26 | 27 | 28 | 29 | ||
| 30 | 31 | |||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ヴェロキサウルス (ノアサウルス類の足)
ヴェロキサウルス・ウニクスVelocisaurus unicusは、白亜紀後期サントニアンにアルゼンチン・パタゴニアのネウケン州に生息した小型のノアサウルス類で、Bonaparte (1991) によって記載された。Bonaparte (1991)は、ケラトサウリアの中にヴェロキサウルス科を設けた。後に複数の研究者によって、ヴェロキサウルスはノアサウルス科に近縁と考えられるようになった。
ヴェロキサウルスのホロタイプ標本は、ほとんど完全な右後肢(大腿骨はない)である。その後、同じ特徴をもつ第2標本が発見されたので、Brissón Egli et al. (2016)によって記載されている。第2標本には大腿骨、脛骨、中足骨、趾骨が含まれているが、中足骨はホロタイプよりも不完全のようである。
ヴェロキサウルスは、後肢だけのノアサウルス類である。どうして後肢だけでノアサウルス類と判定できるのか。というよりも、これを見ればノアサウルス類の後肢の特徴がわかるはずである。
Brissón Egli et al. (2016) によるとヴェロキサウルスは、以下の形質の組み合わせによって識別される。
1)大腿骨の近位端の断面が強く亜三角形で、外側面と内側面が前方で合流して厚い稜crestをなしている。
2)脛骨はコエルロサウルス類のように細長く、その遠位端の前面が平坦で、高く幅広い距骨の上行突起と結合するための広い面となっている。
3)第IIと第IV中足骨は棹状rod-likeで、他の小型のアベリサウロイドの板状laminarの中足骨と異なる。
4)趾骨IV-1は前後に短く背腹に高く、背側部分が側扁していて内側にカーブしている。
1)大腿骨について:カルノタウルスのような多くのアベリサウロイドでは、他の獣脚類と同様に大腿骨の断面は卵形である。マシアカサウルスでは、大腿骨の近位半分の断面が亜三角形で、その頂点が前方を向いている。Evans et al. (2014) はモロッコのケムケム産の分離した大腿骨をノアサウルス科としているが、この大腿骨も近位の断面が亜三角形である。この点でヴェロキサウルスはマシアカサウルスとモロッコの標本によく似ている。しかしヴェロキサウルスでは、前方の頂点が非常に厚く、長軸方向に伸びた稜をなす点がユニークであるという。
2)脛骨について:ピクノネモサウルス、インドスクス、マジュンガサウルスのようないくつかのアベリサウロイドでは、脛骨の遠位端に距骨の上行突起と関節するための斜めの稜がある。これは基盤的テタヌラ類と共有する原始形質であるという。アウカサウルスのような他のいくつかのアベリサウロイドとヴェロキサウルスでは、距骨の上行突起が幅広く、遠位端の前面のほとんどを占めていたと考えられる。これはコエルロサウルス類にみられる状態である。ヴェロキサウルスでは、距骨の上行突起そのものは保存されていない。しかし遠位端の前面は顕著に扁平で、距骨の上行突起のための非常に丈の高い三角形の関節面となっている。Bonaparte (1991) も非常に大きい三角形の上行突起を復元している。このような形態は他のアベリサウロイドにはみられないもので、デイノニコサウリアやオルニトミムス類のような派生的なコエルロサウルス類のものとよく似ている。(アウカサウルスでは上行突起が四角形)
3)中足骨について:Bonaparte (1991) はヴェロキサウルスの特徴として、中足骨の独特のプロポーションをあげている。ヴェロキサウルスでは第IIと第IV中足骨が細くなり、第III中足骨の幅が第IIと第IV中足骨の幅の3倍以上ある。Bonaparte (1991)はこの独特の中足骨の形態は、おそらく現生のレア科にみられるような極端な走行性と関連していると考えた。一方Carrano and Sampson (2008) は、この形質はノアサウルス科に広くみられるとした。実際、エラフロサウルス、マシアカサウルス、ノアサウルスは横方向に圧縮された板状laminarの第II中足骨をもつ。しかしヴェロキサウルスでは、第II中足骨の軸は棹状rod-likeで、断面が亜卵形subovalであるという。それがヴェロキサウルスの固有の形質というわけである。
まあ第III中足骨に対して第IIと第IVが細くなっている、と捉えれば、ノアサウルス科に共通しているとはいえるのだろう。
この幅の狭い第II中足骨については、系統解析に関連してさらに詳しく論じている。
元々、ノアサウルスの原記載で、ノアサウルスを他の軽快な獣脚類と区別する特徴が、この幅の狭い第II中足骨であった。その後Carrano et al. (2002) (マシアカサウルスの記載)はこの形質を、ノアサウルスとマシアカサウルスを結びつけるノアサウルス科の共有派生形質と考えた。これがその後の多くの系統研究に用いられている。
しかしラヒオリサウルス、スコルピオヴェナトル、アウカサウルスのようないくつかのアベリサウルス類では、第II中足骨の近位半分の幅が狭くなり、板状に近づいており、ノアサウルスの状態と似ているという。そのためNovas (2009)は、幅の狭い第II中足骨はアベリサウルス上科全体の特徴かもしれないと考えている。一方でマジュンガサウルスやラジャサウルスのような一部のアベリサウルス類では、第II中足骨の近位端は広く、全く狭くなっていない。この形質はアベリサウルス上科の中で従来考えられたよりも複雑な分布をしているかもしれないと述べている。なるほどアベリサウルス科の中でも、スレンダー型とがっしり型では異なる傾向を示すというのは興味深い。
Brissón Egli et al. (2016)は、ヴェロキサウルスとノアサウルスやマシアカサウルスとでは断面の形状が異なることから、この形質をノアサウルス科の共有派生形質とみなすことには注意が必要であるといっている。
しかし前述のように、エラフロサウルスの論文の系統解析では、ヴェロキサウルスはノアサウルスやマシアカサウルスと同様に幅の狭い第II中足骨をもつとしている。板状とか棹状といっても定量的に定義しないと仕方がないので、数値で分けたようである。
参考文献
Federico Brissón Egli, Federico L. Agnolin & Fernando Novas (2016) A new specimen of Velocisaurus unicus (Theropoda, Abelisauroidea) from the Paso Córdoba locality (Santonian), Río Negro, Argentina, Journal of Vertebrate Paleontology, 36:4, e1119156, DOI: 10.1080/02724634.2016.1119156
コメント ( 0 ) | Trackback ( 0 )
エラフロサウルス
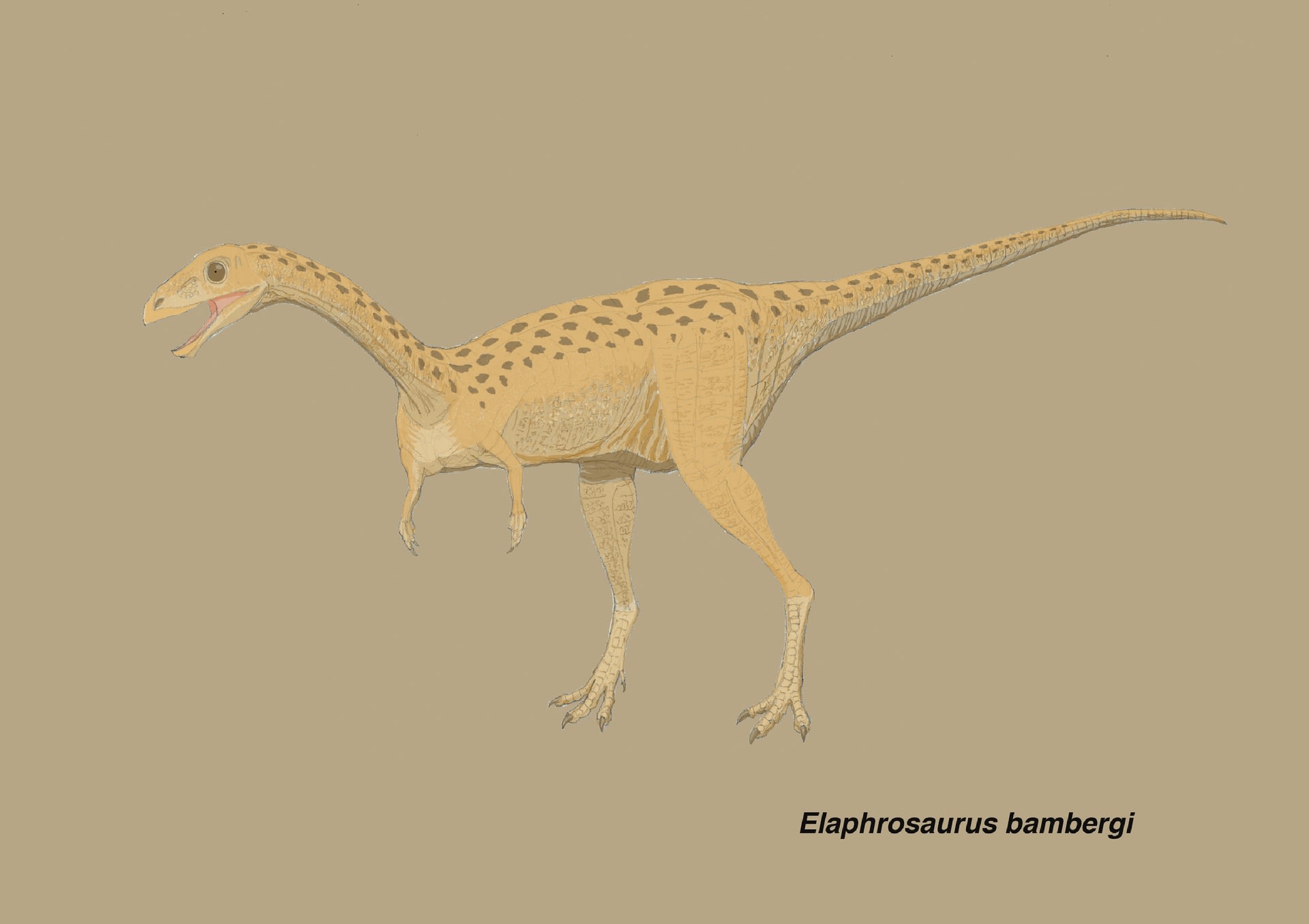
エラフロサウルスは、ジュラ紀後期キンメリッジアンにタンザニア(テンダグル層)に生息したノアサウルス類で、ヤネンシュJanenschによって1920年に命名され1925年に詳細に記載された。ほっそりした体形から、かつてはオルニトミムス類と近縁と考えられていたが、最近の詳細な系統解析により、ケラトサウリアの一員と考えられるようになった。ただしケラトサウリアの中での系統的位置については、研究者によってまちまちで、最近まで定まっていなかった。
エラフロサウルスのホロタイプ標本は、組み立てられて1929年以来、ベルリンの自然史博物館で展示されてきたため、再研究ができなかったが、最近の博物館の展示改装の際に、完全に解体されることになった。そこで、この機会にRauhut and Carrano (2016) はエラフロサウルスの再記載と系統解析を行った。
推定全長6 mでオルニトミムス類のような体形であり、系統的にはリムサウルスと最も近縁なので、ここでは歯のないクチバシで植物食と推測した。前肢は断片的にしか見つかっていない。一言でいうとケラトサウリアの中で、オルニトミムス類のように速く走れる植物食に進化したものと思われる。つまりケラトサウルスやアロサウロイドのような捕食者に襲われる側であろう。
エラフロサウルスのホロタイプ標本は、胴体の部分骨格で頭骨はない。Janensch (1920, 1925)が記載したオリジナルのホロタイプは、16個の仙前椎、6個の仙椎、18個の尾椎、中央の尾椎の血道弓、左の上腕骨、左右の腸骨、左の恥骨、左右の座骨、左の大腿骨、脛骨、腓骨、距骨‐踵骨、中足骨II, III, とIVの基部、3個の趾骨である。その後Janensch (1929)は、左右の肩甲烏口骨、右の中手骨II、左の中手骨IVを追加で記載した。これらは同じ発掘地から発見され同一個体と考えられるので、今回の論文でもホロタイプに含まれている。一方、Janenschは多数の歯をエラフロサウルスに含めているが、頭骨が見つかっていない以上、これらの歯はエラフロサウルスとは認められない。
エラフロサウルスの特徴は、本文中には厳密な標徴形質の記述があるが、論文の要旨ではもっと分かりやすく丸めた表現で書いてある。それは、非常に長く中央がくびれた頸椎、幅広い肩帯と特殊化した前肢、比較的小さい腸骨、細長い後肢と非常に小さい距骨の上行突起、である。
他のノアサウルス類とも区別されるエラフロサウルスに固有の形質は、頸椎の後部に顕著なventrolateral lamina がある;頸椎の前関節突起と後関節突起が細長く、長さが幅の1.5倍以上ある;頸椎にエピポフィシスがない;第2中手骨の遠位端が腹側に曲がっていて明確な段stepがある;距骨の上行突起が非常に小さい、などである。
その他に、形質の固有の組み合わせとして、肩甲骨が幅広く幅が長さの20%以上;肩甲骨の基部で関節窩の上に深い窪みがある;腸骨のbrevis shelf が側方に拡がっている、などの形質が多数あげられている。これらの中にはノアサウルス類に共通の形質も含まれているようだ。

頸椎と胴椎はすべて前後に長いが、特に頸椎は長く伸びている。中央の頸椎では、椎体の長さが高さの4倍以上あり、これはコエロフィシス類やオルニトミムス類と同様である。また頸椎の椎体は、中央で背腹に強くくびれているだけでなく、左右にもくびれている。椎体の側面は前後2対の含気窩によってえぐられているので、本来の椎体は正中線上の壁のような部分だけとなっている。その椎体の腹側面は平面的でキールはないが、エラフロサウルスでは腹側面の後半部は左右のventrolateral laminaの間で凹んだ形となっている。リムサウルス、マシアカサウルス、カルノタウルス、マジュンガサウルスなど多くのケラトサウルス類でも椎体の腹側面は平面的で、側面との間は稜になっているが、エラフロサウルスのように腹外側に突出したフランジ状になっているものはない。
頸椎の前関節突起の最も著しい特徴は非常に細長いことで、神経管孔の幅の70%よりも細い。ディロフォサウルス、ケラトサウルス、アベリサウルス類、アロサウロイドなど多くの獣脚類では、頸椎の前関節突起は長さよりも幅が大きい。ノアサウルス、マシアカサウルス、シギルマッササウルス、オルニトミムス類は比較的細長い前関節突起をもつが、これらもエラフロサウルスには及ばない。
それに対応して後関節突起も細長い形をしている。また、ほとんどの非鳥型獣脚類と異なり、後関節突起の上にエピポフィシスが存在しない。多くのアベリサウロイドでは頸椎のエピポフィシスが強く発達している。
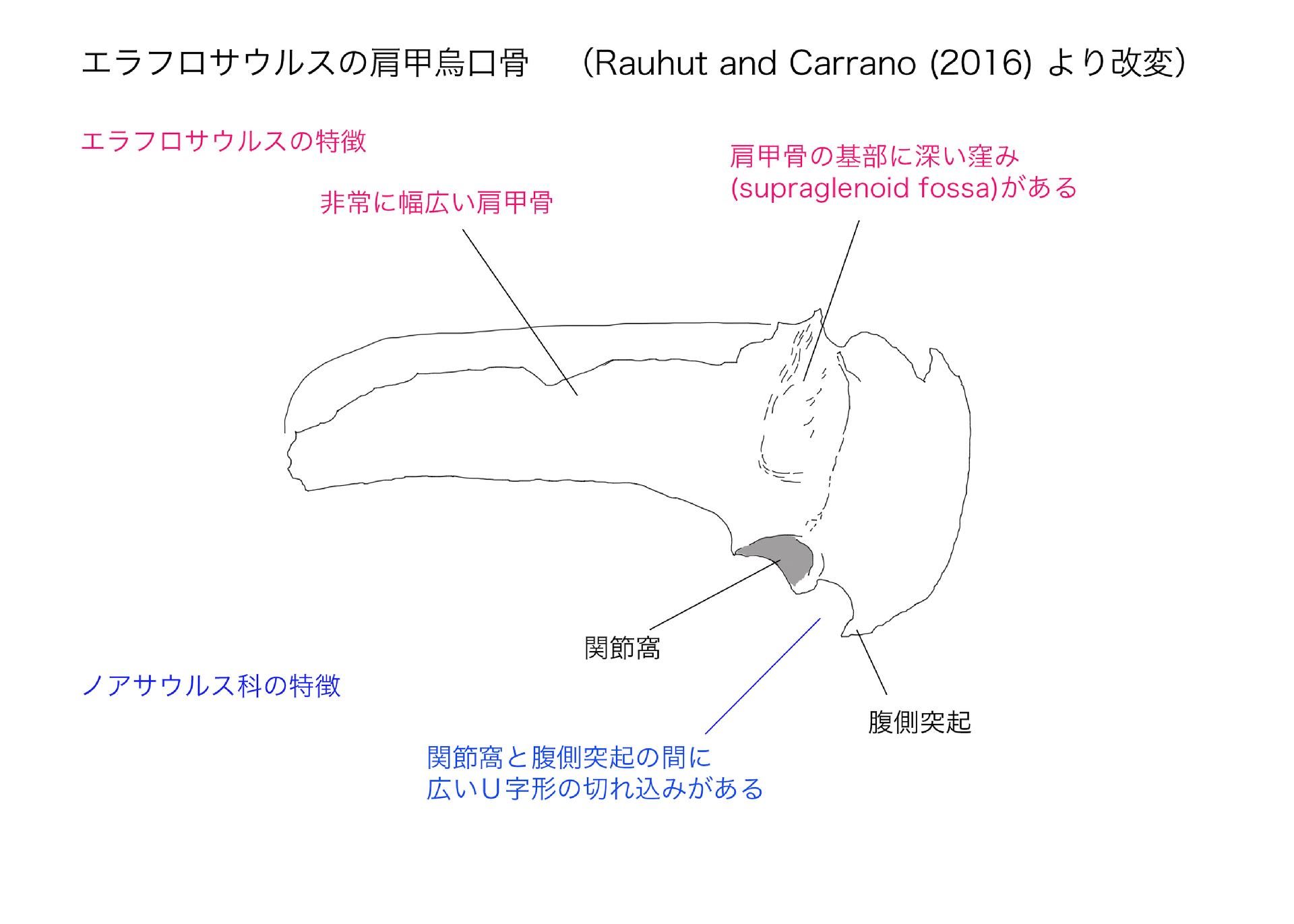
肩帯は左右とも不完全であるが、保存された部分を比較すると肩甲骨は非常に幅広く、幅が長さの25%以上もあると推定される。これは多くの獣脚類の肩甲骨が20%以下であるのと対照的である。珍しい特徴として、肩甲骨の基部(烏口骨の近く)で関節窩の上にはっきりした窪みsupraglenoid fossa がある。同じような窪みは、他にはカルノタウルスとマシアカサウルスにのみみられるが、これらではエラフロサウルスほど発達していない。
エラフロサウルスの前肢は上腕骨と2個の中手骨しか見つかっていない。上腕骨は細長く、三角筋稜は非常に小さく縮小している。獣脚類では珍しく上腕骨頭が内側に傾いているが、この特徴はマシアカサウルスとアルゼンチンで発見されたアベリサウロイドの上腕骨にみられる。
Janenschは2個の中手骨をIと IVとしたが、大きい方の骨は遠位の関節面が対称形であることからIとは考えにくい。この骨はリムサウルスの第III中手骨と似ているが、全体の形は多くの獣脚類の第II中手骨と近いことから第II中手骨と同定された。背側の伸筋溝のあたりが窪みになっており、その端にはっきりした段stepがある。
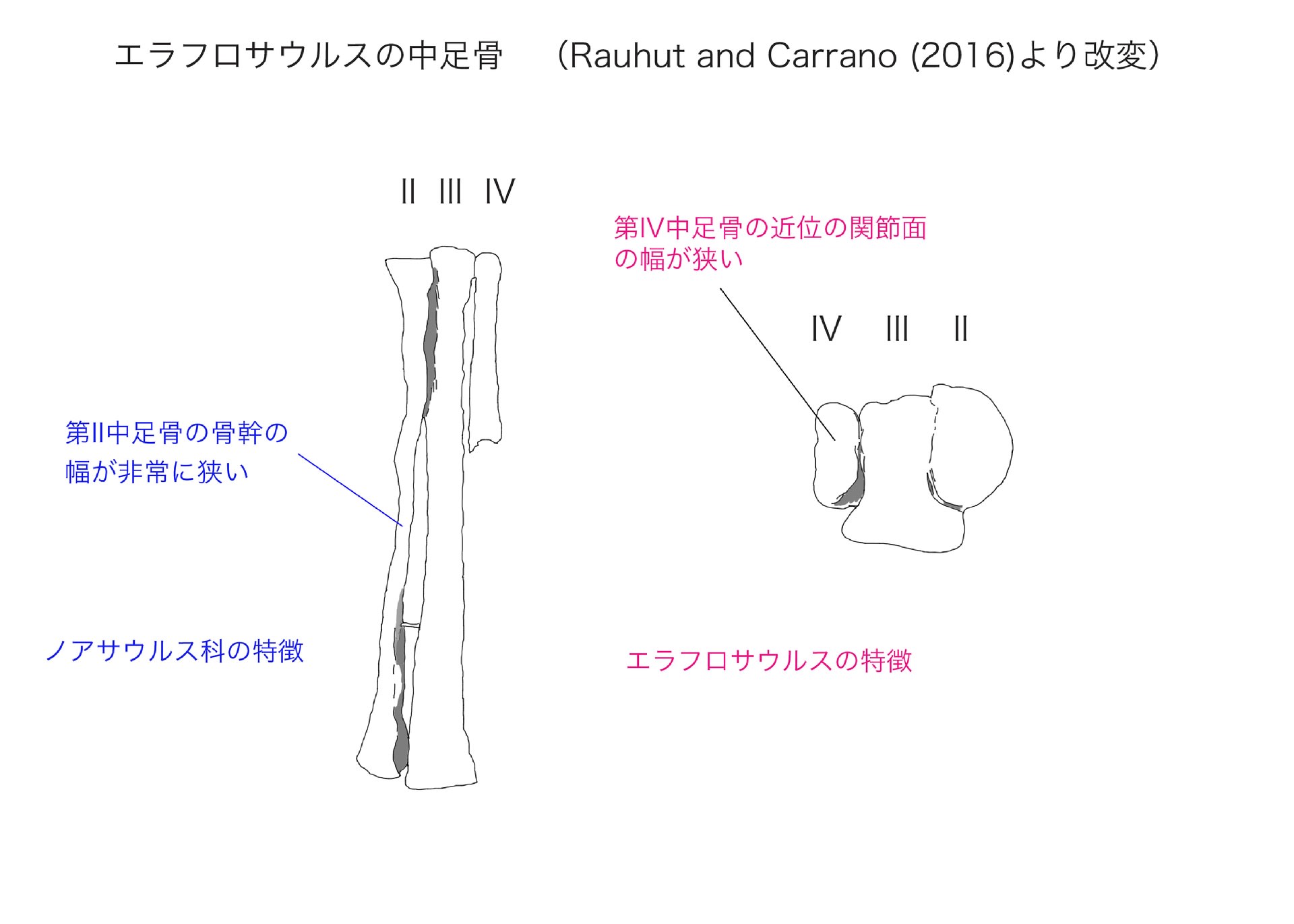
関節した中足骨II, III, IV は非常に細長い。IIIが最も太く、長いようであり、IIはそれよりもやや短いが、IVの遠位側2/3は保存されていない。第II中足骨は近位の関節面がD字形になっていて第III中足骨と接する面が直線的である。
第II中足骨の軸は第III中足骨に寄り添うように細く、縦に扁平になっていて、前後の長さ(奥行き)が左右の幅のほとんど3倍にもなっている。これはノアサウルス、ヴェロキサウルス、マシアカサウルスと同様である。また第II中足骨の幅は第III中足骨の約40%しかなく、これはヴェロキサウルス、マシアカサウルスと同様である。
第III中足骨の近位の関節面は、後方で内側外側ともに拡がったT字形をしている。これをアンタークトメタターサスantarctometatarsusという。
第IV中足骨の近位の関節面は非常に幅が狭くなっており、前後の長さが左右の幅の2.5倍ある。これは他のどの獣脚類にもみられないエラフロサウルスの固有形質である。ヴェロキサウルスやマシアカサウルスでも第IV中足骨の幅はもっと広い。
Rauhut and Carrano (2016)の系統解析では、ケラトサウリアの中にケラトサウルス科とアベリサウルス上科があり、アベリサウルス上科はアベリサウルス科とノアサウルス科に分かれた。ノアサウルス科の中には2つのクレード、エラフロサウルス亜科とノアサウルス亜科がみとめられた。エラフロサウルス亜科にはエラフロサウルスとリムサウルスが含まれ、ノアサウルス亜科にはノアサウルス、マシアカサウルス、ヴェロキサウルスが含まれた。
参考文献
Rauhut, O. W. M. and Carrano, M. T. (2016) The theropod dinosaur Elaphrosaurus bambergi Janensch, 1920, from the Late Jurassic of Tendaguru, Tanzania. Zoological Journal of the Linnean Society, 178, 546-610.
コメント ( 0 ) | Trackback ( 0 )
マシアカサウルスの成長速度

アベリサウロイド(アベリサウルス上科のメンバー)は白亜紀後期のゴンドワナ地域で優勢な捕食者であり多様なグループであるが、多くの種類は化石が不完全なため、詳しい研究が進んでいない。アベリサウルス上科には大型のアベリサウルス科と小型のノアサウルス科が含まれるが、例えばノアサウルス類がどのように小型化したかなどの進化傾向についてもよくわかっていない。
マダガスカルのMaevarano Formationから発見されたマシアカサウルスは、最もよく保存されたノアサウルス類であり、これまでに大きさの異なる複数の個体の分離した骨が多数見つかっている。これは骨の成長過程の研究に適している。そこで、Lee and O’Connor (2013) は大きさの異なる4本の大腿骨と3本の脛骨を選んで、骨幹中央部の切片を作成し、明視野および偏光光学顕微鏡で組織像を観察した。また個々の標本で成長線の周長を測定し、数理モデルを用いて成長曲線を再構築した。
他の多くの脊椎動物と同様に、マシアカサウルスも限定的成長determinate growthを示している。最も大きい脛骨(UA8685)の切片をみると、外側にexternal fundamental system (EFS) が保存されている。EFSは極端に緩慢な成長しかしない骨組織であり、骨の成長が完了したことを示す組織学的指標とみなされている。EFSの中に2本の成長線LAGがあるので、この個体は体の成長が止まってから少なくとも2年は生存していたことになるという。このことからマシアカサウルスは、(大型の獣脚類の幼体などではなく)比較的小型の状態で成長が止まっていた、小型の獣脚類であることが確認された。
偏光顕微鏡で見ると、マシアカサウルスの大腿骨と脛骨には、平行繊維骨parallel-fibered boneが多くみられる。現生の動物では平行繊維骨parallel-fibered boneは成長が遅い傾向がある。このことから、マシアカサウルスは成長過程を通じてゆっくり成長したと考えられる。成長線LAGの測定と成長曲線の解析から、平均的な個体は、大型犬と同じくらいの大きさに成長するのに8~10年かかったと考えられた。成長速度が最も高いのは3~4才であるが、この時期でさえ大腿骨と脛骨の骨幹中央の周長は1年に約7 mmしか増加していない。
著者らは以前、ワニ(ミシシッピアリゲーター)の骨について研究したことがあるので、今回8個体のアリゲーターの大腿骨の切片について、比較のためマシアカサウルスと同じ手法で解析してみた。その結果、アリゲーターはマシアカサウルスよりも40%遅く成長したことが示唆された。つまりマシアカサウルスの成長は、現生のワニよりは速かった。
しかし、マシアカサウルスの成長速度は、同じような大きさの他の獣脚類とは非常に異なっている。コエロフィシス、リムサウルス、コンコラプトル、ビロノサウルス、ヴェロキラプトルの長骨の組織は、主に繊維層板骨fibrolamellar boneからなっている。繊維層板骨fibrolamellar boneは一般に、平行繊維骨parallel-fibered boneよりも速く形成される骨組織である。さらにコエロフィシスとリムサウルスの成長の予備的な解析からは、最も大きな個体は4~6才であったことがわかっている。コンコラプトル、ビロノサウルス、ヴェロキラプトルについてはデータがないが、組織学的類似性から同じくらいと予想される。もしそうならば、マシアカサウルスの成長速度はこれらの小型獣脚類と比べて約40%遅かったことになる。
この大きな違いの理由は不明であるが、系統学的要因や生態学的要因が考えられる。Maevarano Formationの古生態学的研究からは、この地域が季節によって変動する、半乾燥気候であったとされている。少なくとも乾期には生物資源が不足するような厳しい環境に対する適応として、体の維持コストを下げるために低い成長速度が進化したのかもしれない、としている。
参考文献
Andrew H. Lee & Patrick M. O’Connor (2013) Bone histology confirms determinate growth and small body size in the noasaurid theropod Masiakasaurus knopfleri. Journal of Vertebrate Paleontology, 33:4, 865-876.
コメント ( 0 ) | Trackback ( 0 )
マシアカサウルス

大恐竜展では「ノアサウルス科の一種」が展示されていた。これがなかなか可愛らしくて個人的には気に入ったが、そもそもどういう特徴からノアサウルス科とわかるのであろうか。
ノアサウルス類は白亜紀のゴンドワナ地域に生息した小型の獣脚類で、アベリサウルス類と近縁な関係にある。アベリサウルス科とノアサウルス科を含むより大きなグループをアベリサウルス上科Abelisauroideaと称する。従って、アベリサウルス類の祖先がどんな動物だったか考える上で興味深い。例えば、「ノアサウルス科の一種」の復元キャストでは前肢がかなり短かったが、ノアサウルス類の前肢はどの程度見つかっているのだろうか。またこの復元キャストでは、推定で作ったと思われる頭部の下顎が、ちょっとマシアカ風に反っていたのも気になる。下顎が反っているのはマシアカサウルスに固有の特徴だと思うが、他のノアサウルス類の下顎は発見されているのだろうか。
マシアカサウルスは白亜紀後期にマダガスカルに生息した小型の獣脚類で、上顎骨、歯骨、板状骨、角骨(つまり頭骨は上顎と下顎の骨だけ)、頸椎、胴椎、仙椎、尾椎、上腕骨、指骨(撓骨、尺骨などはない)、恥骨、大腿骨、脛骨、脛骨と癒合した近位足根骨(距骨と踵骨)、中足骨、趾骨(末節骨も含む)が見つかっている。
マシアカサウルスは下顎の先端と歯が下方に曲がっていることで有名である。固有の形質としては、前端の4本の歯骨歯が前方に倒れており(procumbent)、最も前端の歯はほとんど水平方向を向いていて、その歯槽は歯骨の腹側縁よりも下がった位置にある。また歯骨歯は高度に異歯性(heterodont)で、前方の歯はスプーン型に近い形で長く、先端が尖ってカギ型に曲がっている。より後方の歯は、普通の獣脚類と同様に扁平で後方にカーブし、前縁と後縁に鋸歯がある。
下顎があまりにもユニークなせいか、それ以外の特徴が書かれた文章をみたことがない。どこがノアサウルス科なのだろうか。ノアサウルス類の化石は断片的で、その特徴とされる形質は文献によって少しずつ異なるが、一つのポイントは「頸椎の神経弓」らしい。
マシアカサウルスの頸椎では、神経棘突起が小さく、椎体の前半部の上にある。また後関節突起とエピポフィシスが強く後方に伸びている(神経棘突起から後関節突起までの距離が、神経棘突起から前関節突起までの距離の2倍くらいある)。エピポフィシス自体は小さく後関節突起から後方に張り出してはいないという。マシアカサウルスの記載論文(Carrano et al., 2002)によると、アベリサウルス上科の中で、マシアカサウルス、ノアサウルス、ラエヴィスクスがノアサウルス科としてまとめられる。これら3属はいずれも神経棘突起が小さく前方にあり、後関節突起が強く後方に伸びている。
マシアカサウルスとノアサウルスの2属は、その他に上顎骨と中足骨の形質を共有している。上顎骨の前眼窩窩の腹側縁がはっきりと(土手のように)盛り上がっていること、上顎骨の口蓋突起が単純であること、第2中足骨の軸が左右に薄くなっていることである。
マシアカサウルスの上顎骨には7個の歯槽が保存されているが、せいぜいあと2、3個しかないらしいという。ノアサウルスも10~11個と考えられているので、上顎骨の歯の数が少ない点も共通している。前眼窩窩の腹側縁が盛り上がっていることはコエロフィシス類にもみられるが、コエロフィシス類の上顎骨では歯の数が多く、前上顎骨と上顎骨の間にくびれsubnarial gap (notch)がある。
またマシアカサウルスの上顎骨では萌出した歯は保存されていないが、歯槽の角度をみると、前方の歯はやはり前に傾いていたことがわかるという。最も前方の歯槽は大体、前上顎骨との結合面と平行で、水平から40°というからかなり斜めになっている。歯槽の角度は後方にいくにつれて次第に垂直に近づき、5番目の歯槽で完全に垂直になるという。そして最も前方の歯槽は断面が円形で、後方の歯槽は扁平(楕円形)になる。つまり、マシアカサウルスの歯は、下顎だけではなく上顎の上顎骨でもprocumbentでheterodontである。前方の2、3本はある程度傾いてみえると思われる。なお前上顎骨は発見されていないので、その歯は推定で前方に傾いている(出っ歯)ように復元されている。2005年の科博での復元キャストでは前上顎骨の歯は傾いているが、上顎骨の歯は斜めになっていないようにみえる。歯槽と歯冠ではまた角度が違ってくるとしても、もう少し傾いているように造ってもいいのではないだろうか。
下顎の歯骨では、最前端の歯がほとんど水平(10°)で、2番目、3番目は20°、35°で、かつ側方に10°開いている。また、歯骨の後端の形態から外側下顎窓が非常に大きいことがわかるが、これはアベリサウルス類と似ている。
上顎、下顎とも、後方の歯は普通の獣脚類と同じようなナイフ型をしている。Carrano et al. (2002) は、前方の歯が非常に特殊化しているだけでなく、そのわりに後方の歯が普通であることも不思議であると述べている。前方の歯で小型の動物を捕らえ、後方の歯で切断したのかもしれないという。
白亜紀後期にアルゼンチンに生息したノアサウルス・レアリは、1980年に記載された時にはコエルロサウルス類と思われていた。模式標本は断片的で、上顎骨、方形骨、鱗状骨、頸椎の神経弓、胴椎の椎体、頸肋骨、第2中足骨、趾骨、末節骨が発見されている。下顎や前肢の骨は見つかっていない。
参考文献
Bonaparte, J. F. and J. E. Powell. 1980. A continental assemblage of tetrapods from the Upper Cretaceous beds of El Brete, northwestern Argentina (Sauropoda-Coelurosauria-Carnosauria-Aves). Me'moires de la Socie'te' Ge'ologique de France, Nouvelle Se'rie 139:19-28.
Carrano, M. T., S. D. Sampson, and C. A. Forster. 2002. The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology 22: 510-534.
コメント ( 3 ) | Trackback ( 0 )