獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2024年12月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | 31 | ||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |

カルノサウリアは復活するか (補足)

さきのアスファルトヴェナトルの記事に書いてあるが、よくわからない人もいるかもしれない。ある程度、恐竜の分類や研究史を知っている方なら「カルノサウリアの復活」といえばピンとくるのであるが、初心者の方にはさっぱりわからない方もおられるだろう。
私はサイエンスライターでもないし、わかりやすく解説する適役でもないが、たまには噛み砕いて説明してみようと思う。
昔(結構むかし)
恐竜研究の初期には、獣脚類のうち大型肉食恐竜をカルノサウリア(カルノサウルス類)、小型の肉食恐竜をコエルロサウリア(コエルロサウルス類)と分類していた。そのため、現在は別々の系統であるケラトサウルスも、メガロサウルスも、アロサウルスも、ティラノサウルスも、みんな一緒くたにされていた。カルノタウルスでさえ、発見の初期にはカルノサウリアと称されていたくらいである。
現代(分岐分類の普及から現在、図のA)
その後、恐竜の分類研究が進み、多数の形質を用いた分岐分析で系統研究が行われるようになると、それまでのカルノサウリアにはどうも異なる系統が含まれていたことがわかってきた。ケラトサウルスはケラトサウリアという、かなり根本から異なる系統に属し、他の多くの肉食恐竜はテタヌラ類という大きなグループに含まれた。テタヌラ類の中ではメガロサウルスの仲間が最も原始的な部類で、メガロサウルス類とよばれる。メガロサウルス類とスピノサウルス類、あとピアトニツキサウルスなど若干の種類を含めてメガロサウルス上科(メガロサウロイデア)といい、これがテタヌラ類の中で最も基盤的なグループである。(注1)
より進化的なテタヌラ類がアヴェテロポーダで、これはアロサウルス上科(アロサウロイデア)とコエルロサウルス類(コエルロサウリア)に分かれる。アロサウルス上科(アロサウロイデア)にはメトリアカントサウルス科、アロサウルス科、カルカロドントサウリア(カルカロドントサウルス科とネオヴェナトル科)が含まれた。
一方のコエルロサウルス類のうち、基盤的なところからティラノサウルス上科(ティラノサウロイデア)が分岐し、その中のティラノサウルス科が白亜紀末の大型捕食者を生み出した。
ということで「カルノサウリア」は昔のよくわかっていなかった時代の概念で、今では肉食恐竜の代表のように思われるティラノサウルスは、実は鳥に近い小型のコエルロサウルス類の一員なんだよ、というのが、よく恐竜本などに説明されていることである。単純化していえば、アロサウルスとティラノサウルスは、原始的なメガロサウルスと比べれば互いに近縁ということだった。
もう一点、最近はカルノサウリアという用語自体があまり使われなくなっていた。「カルノサウルス類」と「(広義の)アロサウルス類」はどう違うの?ということでモヤモヤしている方もいるだろう。多くの場合、(テタヌラ類が確立した後の)カルノサウリアとアロサウロイデアの内容は同じだったが(カルノサウリアの中にアロサウロイデア以外の種類を含める考えもある)、昔の混とんとした時代の「カルノサウリア」との混同を嫌って、カルノサウリアは用いないでアロサウロイデアが用いられるようになった。最近の獣脚類の文献ではcarnosaurs (カルノサウルス類)はあまり用いられず、より明確にallosauroids (アロサウロイド)と表記されることが多い。このブログで常にアロサウロイドと書いているのはそのためである。メガロサウルス上科の動物はメガロサウロイドmegalosauroids とするか、基盤的テタヌラ類 basal tetanurans と括られるので、カルノサウリアを使う必要はないわけである。
つい最近の一部の研究 (B)
2012年のスキウルミムスの研究で、幼体ではテタヌラ類の各系統を分類するのが困難であることと、メガロサウルス上科とアロサウルス上科を合わせて、コエルロサウルス類は含まない「カルノサウリア」の可能性が浮上した。そして2019年、アスファルトヴェナトルの発見により、再び「カルノサウリア」の可能性が指摘されたことになる。
ここで注意すべきこととして、「カルノサウリア」の復活といっても、昔のようにケラトサウルスやティラノサウルスが戻るわけではない。コエルロサウルス類が根元から分かれていて、メガロサウルス上科とアロサウルス上科がより近縁になったということである。つまり単純化すると、ティラノサウルスに対してメガロサウルスとアロサウルスがより近縁になった。
さらにアスファルトヴェナトルの解析結果では、メガロサウルス上科は単系でなく、アロサウルス上科へと向かう系統の側枝である可能性も出てきたということである。
もちろん、「つい最近こうなった」とか、確定したという話ではない。可能性があるということである。しかしスキウルミムスは幼体なのでどうかなーと思われたが、アスファルトヴェナトルは成体でアロサウロイドとメガロサウロイドの中間的なものが出てきたということで、意外と可能性があるのではないかと。
結局、大してわかりやすくなく、難しい説明になった。
注1:ここは簡略化して説明しているので、不正確だと突っ込まないように。厳密にいうと最も基盤的なテタヌラ類は、メガロサウロイデアにも属さない。ここはメガロサウロイデアとアヴェテロポーダを合わせたオリオニデスの話をしている。
コメント ( 0 ) | Trackback ( 0 )
アスファルトヴェナトル・ヴィアリダディ

(解説は前の記事参照)
なんだかんだいってもアルゼンチンはすごい所で、アベリサウルス類もまるっと出るし、メガラプトル類も次々に発見されている。そしてピアトニツキサウルスよりも完全な、こんなものがほぼ全身骨格で見つかるとは。
にせアロサウルスの見分け方
かなりアロサウルスと似た動物で、体形も大まかには同様である。頭骨図を見ると眼窩の幅が狭く縦長のようにも見えるが、写真を見ると必ずしも言い切れないかもしれない。
外観上確実なのは、後眼窩骨にちょっと突起がある、下顎の歯骨の先端が微妙にふくらんでいる、手の第3指が小さい、といった所だろうか。
コメント ( 0 ) | Trackback ( 0 )
アスファルトヴェナトルとテタヌラ類の系統関係
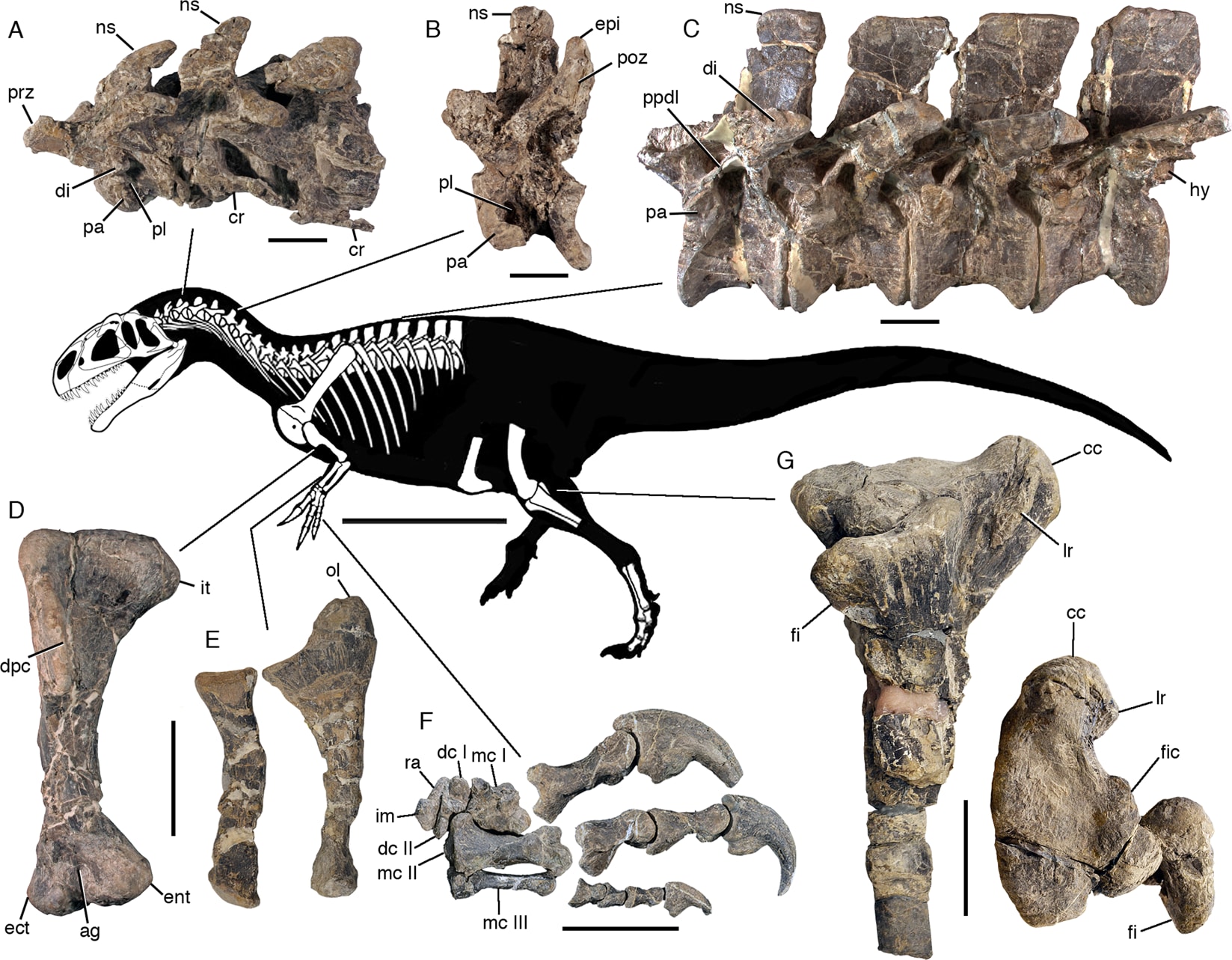
Copyright 2019 Rauhut and Pol
アスファルトヴェナトルは、中期ジュラ紀の初めごろ(前期ジュラ紀の終わりトアルシアンから中期ジュラ紀のバジョシアン、Canadon Asfalto Formation)に、アルゼンチンのチューブート州セロ・コンドルCerro Condor村に生息したテタヌラ類で、おそらく最古のアロサウロイドと考えられている。Rauhut and Pol (2019) により記載された。
テタヌラ類はメガロサウルスから鳥まで含む最も繁栄した獣脚類のグループである。最古のテタヌラ類は中期ジュラ紀の初期に出現していたと考えられるが、その頃のテタヌラ類の化石記録はほとんどが断片的なもので、テタヌラ類の初期進化についてはよくわかっていない。近年の系統解析ではテタヌラ類の中に、3つの主要なグループを置いている。メガロサウルス上科(メガロサウルス科とスピノサウルス科)、アロサウルス上科(メトリアカントサウルス科、アロサウルス科、カルカロドントサウリア)、コエルロサウルス類である。この3つの系統関係について、また個々の種類の位置付けについては現在でも議論があり、確定していない。
(メガロサウルス上科のメンバーをメガロサウロイド、アロサウルス上科のメンバーをアロサウロイドと呼ぶ。)
アスファルトヴェナトルのホロタイプ標本は、この時代のテタヌラ類として最も完全なもので、分離した完全な頭骨と胴体の部分骨格からなる。前半身すべてと部分的な恥骨、後肢がある。全体にアロサウルスと似ており、頭骨が75-80 cm、推定全長は7-8 m である。
頭骨はアロサウルスとよく似て、涙骨の突起もあるし上顎のラインはまっすぐで頰骨の突起まである。アロサウルスの直系の祖先といわれれば納得するくらい似ている。さらに、下顎にはアロサウルスに固有の特徴とされていたantarticularという骨がある。
アスファルトヴェナトルの特徴は、以下の形質の組み合わせからなる。前上顎骨歯にはよく発達した後縁の鋸歯と非常に細かい前縁の鋸歯をもつ;後眼窩骨の背側に小さな角状突起がある;傍後頭骨突起と大後頭孔の間の外後頭骨に、顕著な水平の稜がある;下顎に骨化したantarticular がある;platycoelousな頚椎;第3と第4頚椎の神経棘が三角形で後方に傾いている;後方の頚椎に腹側のキールがなく、前方の胴椎にもほとんど発達していない;関節した中手骨部分は長さよりも幅が大きい;手の第3指は第1指、第2指と比べて顕著に小さい。などである。
最近の基盤的テタヌラ類の系統解析では通常、メガロサウルス上科がまず分岐し、アロサウルス上科とコエルロサウルス類が姉妹群をなすようになっている。ところがアスファルトヴェナトルには、従来メガロサウルス上科の共有派生形質と考えられた特徴と、アロサウルス上科の共有派生形質とされる特徴、さらには原始的なテタヌラ類の形質が、モザイク的に混じっている。
メガロサウロイドの特徴としては、上顎骨の上行突起に顕著な屈曲kinkがあること、内側が閉じた(貫通していない)maxillary fenestra 、後眼窩骨の腹方突起の後側に深い溝がある、後頭顆の下に幅広い窪みがある、歯骨の前端が拡がっている、などがある。
アロサウロイドの特徴としては、顕著なsupranarial fossa 、前眼窩窩に鼻骨が参加している、鼻骨に含気孔がある、鼻骨の側方のトサカcrestがある、区分されたlacrimal fenestra 、傍後頭骨突起が強く腹側方を向いている、などがある。
原始的なテタヌラ類の特徴としては、上顎骨の内側に含気性の窪みがない、頚椎の前方の関節面が平らである、肩甲骨が幅広い、がある。
さらにアスファルトヴェナトルは、これまで別の種の固有形質と考えられていた形質を併せもつ。外翼状骨の頰骨突起にある突起(ドゥブレウイロサウルスの固有形質)、第3頚椎の低く後方に傾いた神経棘(アフロヴェナトルの固有形質)、骨化したantarticular (アロサウルスの固有形質)である。
アスファルトヴェナトルを含めた系統解析の結果、基盤的なテタヌラ類の系統関係に大きな変化が生じた。まずメガロサウロイドとアロサウロイドが単系のカルノサウリアをなし、コエルロサウルス類はその外側にきた。またメガロサウルス上科のいくつかのグループは、単系のアロサウルス上科に対して順次外群となった(メガロサウルス上科は多系群となった)。つまりアロサウルス上科へと向かう系統の中から、スピノサウルス科、メガロサウルス科、ピアトニツキサウルス科の順に次々と分岐した形となった。アスファルトヴェナトルは特にアロサウルスと近縁ではなく、ピアトニツキサウルス科とメトリアカントサウルス科の中間段階という、微妙な位置にきている。つまりここではピアトニツキサウルスよりは派生的で、シンラプトルより基盤的ということになる。
ただしテタヌラ類の初期進化・放散の時期には、異なる系統の間で非常に多くの平行進化や収斂が生じているので、この系統関係を正確に推定するのは現在のところ非常に困難であると言っている。伝統的なメガロサウルス上科を保つ分岐図も、確率は少ししか違わないという。そのことが論文のタイトルにも表れている。
いずれにしても世界的に分布していたメガロサウルス段階の原始的なテタヌラ類の中から、同時多発的にアロサウロイド的なものが進化してきたのだろう。そこで個人的に思い出すのは中国の状況で、アロサウロイドに入るか入らないか微妙なものもいる。四川省のレシャンサウルスは最初中国の研究者によってシンラプトル科とされたが、後に欧米の大規模な研究(Carrano et al. 2012)ではメガロサウルス類とされた。しかし中国の研究でもそれなりにアロサウロイドたる根拠を示しているのだから、やはりモザイク的な種類なのではないだろうか。現在どうなっているのかわからないが、アジアのメトリアカントサウルス類(シンラプトル類)はやはりアジアの原始的テタヌラ類の中から進化してきたと考えたい気持ちはある。
参考文献
Rauhut, O.W.M., Pol, D. Probable basal allosauroid from the early Middle Jurassic Cañadón Asfalto Formation of Argentina highlights phylogenetic uncertainty in tetanuran theropod dinosaurs. Sci Rep 9, 18826 (2019). https://doi.org/10.1038/s41598-019-53672-7
コメント ( 0 ) | Trackback ( 0 )
アクロカントサウルス1

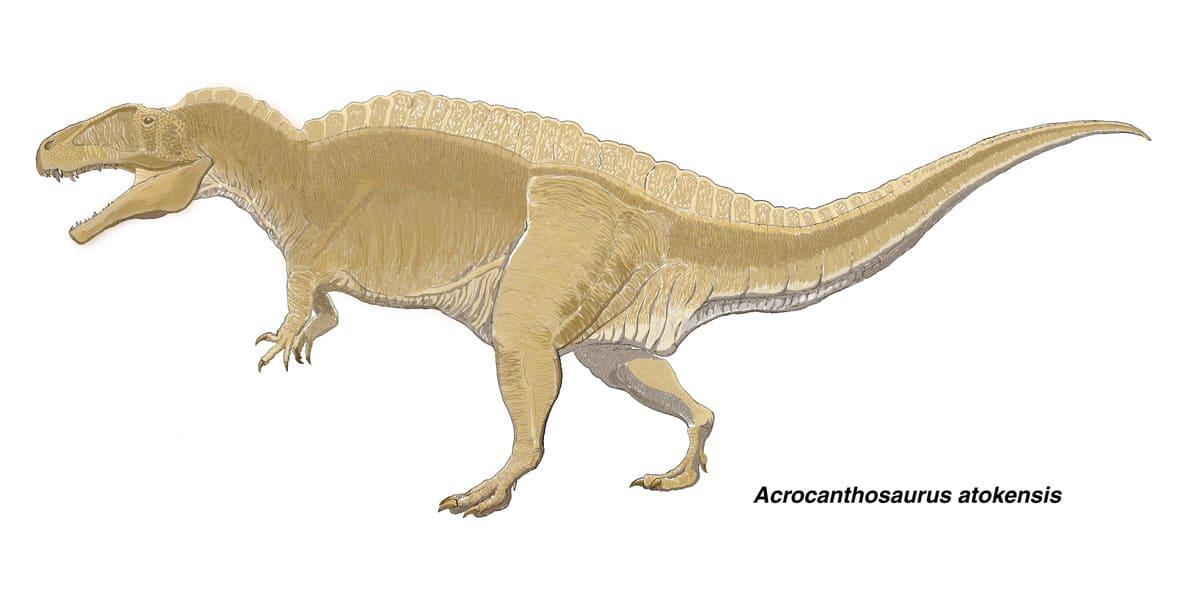
アクロカントサウルスは、白亜紀前期アプト期に北アメリカに生息した大型のカルノサウルス類である。アロサウルス上科に属することは認められているが、アロサウルス科とするか、カルカロドントサウルス科とするかで研究者の意見が一致していない。
米国オクラホマ州の白亜紀前期の地層から発見された2つの部分骨格に基づいて、1950年に記載された(Stovall and Langston, 1950)。それ以来、アロサウルス科とされてきたが、1996年にセレノはカルカロドントサウルス科とし、また1990年にテキサスで発見された保存のよい骨格の解析から、ハリスもカルカロドントサウルス科とした(Harris, 1998)。その後、再びオクラホマ州のアントラーズ層から、ほとんど完全な頭骨を含む、さらに保存のよい全身骨格が発掘され、最終的にはブラックヒルズの手で復元された。この標本のオリジナルはノースカロライナ州立自然科学博物館にある(NCSM 14345)。この標本を研究したカリーとカーペンターは、カルカロドントサウルス類と共通するとされた多くの形質は大きさ依存的size dependent, size relatedなもので系統関係を反映するものではないとし、その他の特徴からやはりアロサウルスに近縁(アロサウルス科)とした(Currie and Carpenter, 2000)。ところがさらにその後、X線CTを用いてホロタイプの脳函のエンドキャストが作製され、脳の特徴はアロサウルスやシンラプトルよりもカルカロドントサウルスとギガノトサウルスに似ており、やはりカルカロドントサウルス科という説が支持された(Franzosa and Rowe, 2005)。The Dinosauria 2nd editionでもカルカロドントサウルス科に含められている。
最大の特徴である脊椎骨の神経棘は、仙前椎、仙椎、前方の尾椎で椎体の長さの2.5倍以上あるという。頸椎の神経棘の前方に三角の突起があり、ひとつ前の神経棘のへこんだ部分にはまるようになっている。
前眼窩窓は非常に大きく、上顎骨にはMaxillary fenestra とPromaxillary fenestraの両方がある。カルカロドントサウルス類とアベリサウルス類では上顎骨の外側面の大部分に粗面があるが、アクロカントサウルスではアロサウルスなどと同様に粗面は前縁と歯列のすぐ上に限られ、大部分はなめらかである。眼窩は縦に長い。涙骨の背側方部はカルカロドントサウルスやギガノトサウルスと同じような低い稜をなす。涙骨の腹側部(眼窩の前縁)に下眼窩突起がある。後眼窩骨の腹側部(眼窩の後縁)にも下眼窩突起がある(NCSM 14345では、はっきりしていない)。比較的大きな三角形の前前頭骨が後眼窩骨と接して眼窩の上のひさし部分に寄与している。カルカロドントサウルス類、アベリサウルス類、大型のティラノサウルス類と同様に、後眼窩骨の上部は肥厚して顕著な水平の突起をつくっている。アクロカントサウルスの歯はナイフ状であるが、カルカロドントサウルス類の歯ほど薄くはなく、またカルカロドントサウルス類に特徴的なエナメルのしわはない。一方、下顎の前端はギガノトサウルスと同様に角張っている。
カルカロドントサウルス科の特徴として涙骨と後眼窩骨の結合部が広いことが挙げられるが、カリーとカーペンターはこの特徴はsize relatedな形質だろうといっている。カルカロドントサウルス類と同様にアクロカントサウルスにも眼窩の上にひさし状の骨(supraorbital shelf)がある。アクロカントサウルスでは、supraorbital shelfは涙骨、前前頭骨、後眼窩骨からなるのに対して、カルカロドントサウルス科のギガノトサウルスではsupraorbital shelfは涙骨、palpebral、後眼窩骨からなるとしている。つまり骨の構成が異なるといいたいらしい。後眼窩骨の下眼窩突起もある程度size relatedといっている。
前肢はアロサウルスと比べるとかなり小さい。大腿骨に対する前肢の長さの比率は、アクロカントサウルスでは81%であるが、アロサウルスのある標本では126%に達するようである。
2002年の恐竜博の全身復元骨格を見る限り、アロサウルスに近縁としても体型はかなり異なるという印象を受ける。大型化にともなって、頭が大きく前肢は小さく、後肢も比較的短い、全体としてがっしりした重々しい体型となっている。軽快なアロサウルスのように速く走れたとは思えない。もちろん、復元骨格は推定を含むものであり、また亜成体や幼体の時期にはアロサウルスのようなスマートな体型だったかもしれないが。この体型でも竜脚類に追い付くには充分ということなのか、あるいは屍肉食だったのだろうか。
コメント ( 0 ) | Trackback ( 0 )