獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2018年8月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | |||||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| 12 | 13 | 14 | 15 | 16 | 17 | 18 | ||
| 19 | 20 | 21 | 22 | 23 | 24 | 25 | ||
| 26 | 27 | 28 | 29 | 30 | 31 | |||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク





| URLをメールで送信する | |
| (for PC & MOBILE) | |
ネオヴェナトルの神経血管網:敏感な吻はスピノサウルスに限らない
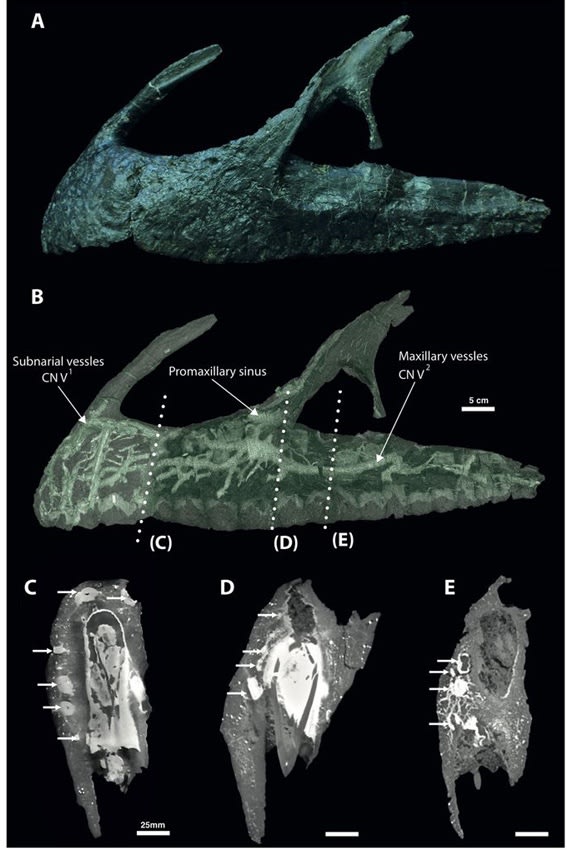
Copyright 2017 Barker et al.
スピノサウルスの半水生適応の研究では、吻部の感覚器の話があった。スピノサウルスの吻(前上顎骨)の表面には多数の孔(神経血管孔)があいており、CTスキャンで内部を観察すると、複雑に枝分かれした神経血管網と思われる管状構造がよく発達している。これは現生のワニと同様に機械受容器と連絡していると考えられ、水中で獲物による振動を感知するなど、半水生適応と関連づけられていた。つまりスピノサウルスのよく発達した神経血管網は、水中で獲物を探すための特殊化と考えられた。バリオニクスについてもそのようにいわれていた。
しかし、本当にスピノサウルス類で発達した形質なのかどうかは、他の獣脚類と比較しないとわからない。実は、スピノサウルス類以外の吻部の神経血管網については、詳細に研究されていなかった。Barker et al. (2017) は、ネオヴェナトルの前上顎骨と上顎骨の表面に、多数の神経血管孔があることに注目して、高精度のCTスキャンで内部構造を研究した。
その結果、ネオヴェナトルの前上顎骨と上顎骨の内部には、スピノサウルスに匹敵するほどよく発達した神経血管網のネットワークが観察された。特に前上顎骨と上顎骨の前半部分で最も発達していて、複雑に枝分かれ、吻合した管状構造がみられた。分岐した枝は最終的に骨表面の神経血管孔に開口していた。
著者らはワニの神経との比較などから、これらの神経血管網を(一部は血管も含まれているが、)主に三叉神経の枝と同定した。前上顎骨には、三叉神経の眼神経CN V1(鼻骨枝の前上顎枝)が分布している。上顎骨には、三叉神経の上顎神経CN V2が分布していて、上顎と歯からの感覚を伝達している。
このようなデータは、吻部の神経の発達に関して、スピノサウルスが獣脚類の中で特別ではないことを示している。典型的な陸上の捕食者であるネオヴェナトルが、スピノサウルスに匹敵する神経血管網と吻部の鋭敏な感覚をもっていることから、これが半水生適応とは考えにくい。ネオヴェナトルが時々、水中の魚などを捕食したことは考えられるが、体の解剖学的特徴などから半水生は支持されない。ネオヴェナトルは他のカルカロドントサウルス類やアロサウロイドと同様に、中型の植物食恐竜を捕食したと考えられている。
スピノサウルスの神経血管網は確かに水中で獲物を検知するのに役立ったかもしれないが、それが水生適応のためだけに発達したというのは言い過ぎであることになる。むしろ、大型の肉食恐竜は一般に、吻部に従来考えられていたよりもよく発達した神経血管網をもっており、獲物の扱いprey handling など他の目的に利用していた可能性が高い。Barker et al. (2017)は3つの目的をあげている。
1つは、肉食恐竜の吻部の鋭敏な感覚は、獲物の解体・処理に用いられたというものである。例えば、骨を避けて肉だけをそぎ取ることが必要になる。ネオヴェナトルの上顎骨歯の表面の微小な摩耗痕が解析されている。そのパターンは、チーターのような骨を噛むことを避ける肉食動物と同じ特徴をもつことから、ネオヴェナトルは骨を避けて肉だけを食べたと推測される。その際に吻部や歯の鋭敏な感覚は、注意深く骨と肉を識別するのに役立ったと考えられる。
2つめは種内のコミュニケーションで、ティラノサウルスなどの大型肉食恐竜は、闘争の際に顔や顎を噛みあうことが知られている。儀式的な闘争などで吻部・顔面の感覚は重要だった可能性がある。
3つめは、巣作り行動の際に穴を掘ったり、適切な温度の場所を探したり、メンテナンスする上でも吻部の鋭敏な感覚は役立っただろうとしている。
確かに吻部の感覚器は、ワニのように水中の感覚に用いるとは限らない。現生鳥類ではキーウィ科、シギ科、トキ科のようにくちばしで餌を探す鳥の吻部に、グランドリー小体やヘルプスト小体という感覚受容器があって、餌を探す際に獲物を認識している。一方オウム類では果実を割るなどの操作に重要であるという。
参考文献
Chris Tijani Barker, Darren Naish, Elis Newham, Orestis L. Katsamenis & Gareth Dyke (2017) Complex neuroanatomy in the rostrum of the Isle of Wight theropod Neovenator salerii. Scientific Reports 7: 3749 DOI:10.1038/s41598-017-03671-3
コメント ( 0 ) | Trackback ( 0 )
特別展「獣脚類 鳥に進化した肉食恐竜たち」(2)

ビセンテナリアは恥ずかしがり屋さんなのか、壁の方を向いている。いや、パネルの説明を読んでいるようにみえる。獣脚類の進化に興味があるのだろう。

今回感動したものの一つが、ズオロンとアオルンである。過去にアオルンの記事は書いているが、これが見られる日が来るとは。以前ズオロンも記事にしようとしたが、結構細かいのであきらめた経緯がある。(「アオルン」の記事参照)

ティラノサウルス類で重要なのはチエンジョウサウルス。今回、キアンゾウではなくチエンジョウと表記されたことはちょっとうれしい(「チエンジョウサウルス」の記事参照)。ローマ字読みするのは自由だが、そもそもquiならスペイン語などのキだが、qiはローマ字読みできないはずである。このチエンジョウサウルスは歯が抜けているためにくちばし状にみえるが、歯を生やすとアリオラムス顔になり、それなりに様になるはずだ。

ティラノサウルス類以上のコエルロサウルス類は、オルニトミムス類、アルバレスサウルス類、テリジノサウルス類、オヴィラプトロサウルス類など、ひと通りそろえた感がある。次の山場はパラヴェス類(近鳥類)で、アンキオルニス、エオシノプテリクス、アウロルニスなどが一堂に会し、エピデクシプテリクスやイーもいる。ジェホロルニス、サペオルニス、孔子鳥類、エナンティオルニス類へと続いていく。

ちょっと注目したのはサペオルニスの歯で、結構立派な歯である。やはり鋭い歯と長い尾があると、獣脚類という気がする。ニワトリのゲノムを元に恐竜を作成するなら、まずは歯と尾椎を目標にするといいのではないか。そうすればラプトルまではできたも同然である。


ミクロラプトル、シノルニトサウルスはともかくティアンユーラプトルもいるとは。さらにウネンラギア類アウストロラプトルとブイトレラプトル。この辺でお腹いっぱいになってきた。
アウストロラプトルは論文の図とは印象が違うが、初めて見るのでそれなりに感慨深い。このキャストは骨の表面のテクスチャーがピアトニツキサウルスと似ているようだが…アルゼンチン製か?これもブイトレラプトル同様、尾椎の関節突起などは未発達となっている。
中国のティアンユーラプトルと並べて、ティアンユーラプトルの前肢が短いことから関連があるかもしれない的なことが説明してある。しかしアウストロラプトルは確かに短いが、ブイトレラプトルやラホナヴィスはむしろ長いわけだから、ウネンラギア類全体に短い傾向があるわけではない。ミクロラプトル類やウネンラギア類を含めてどのグループにも、独立して前肢が縮小する傾向があったのかもしれない。ティアンユーラプトルの論文には、確かにそういった考察があったのを思い出した。(「ティアンユーラプトル」の記事)
今回、近鳥類Paraves 、鳥翼類Avialaeの他、ペンナラプトル類など、専門家の間では定着しているのかもしれないが聞きなれない分類名が、普通に多用されている。恐竜博物館の特別展だからこれでいいのかもしれないが、一般の親子連れのお父さんが「もう少しわかりやすく書けばいいのに」とつぶやくのを耳にした。アンキオルニス科は多分、2017年のカイホンの系統解析あたりかな。
続くかな
コメント ( 1 ) | Trackback ( 0 )
特別展「獣脚類 鳥に進化した肉食恐竜たち」(1)
大好物のご馳走ばかり並べられて、「箸休め」がない。肉料理が続く場合は、前菜やサラダやガーリックトーストもやはり必要である。そんな感じで、最後まで少しも気が抜けないので疲れた。鳥になってしまうとわりと興味を失うので、私がエキサイトしたのは主に前半ではあるが。。
最近は毎年、聖地巡礼しているが、今回はお盆休みに行ったので親子連れなどで混んでいた。館内のレストランは長蛇の列で「獣キャルビ丼」も食べられない。やむなく外のレストランでカレーを食べたが、こちらも相当混雑していた。こちらのショップにも山のような恐竜グッズがある。
今回のテーマは「肉食恐竜から鳥への進化」ではない。鳥への進化だけでなく、獣脚類全体の様々な局面に注目した特別展である。
なぜこういう構成なのか推測すると、福井が所有する獣脚類化石の研究に役立つ標本を集めたということだろう。最近メガラプトル類とされているフクイラプトルの研究をさらに発展させるためには、ネオヴェナトルやメガラプトルと比較することが必要である。奇妙な基盤的マニラプトル類フクイベナートルの位置づけなどを解明するために、原始的なコエルロサウルス類にまでさかのぼって研究する必要があり、そのためにビセンテナリア、ズオロン、アオルンを持ってきた。そして北谷化石鳥の研究には、エオコンフキウソルニスをはじめ白亜紀前期の鳥類が必要になった。そういう感じではないか。

今回はヘレラサウルスやコエロフィシスに時間はかけられない。カルノタウルスは頭をやや下げて突進していた。なにげに見る機会がないのがアベリサウルス頭骨だが、復元したものなので、おそらくこの通りではないようだ(「アベリサウルス類のアロメトリー」の記事)。

ピアトニツキサウルスは基盤的テタヌラ類で、最近はピアトニツキサウルス科(ピアトニツキサウルス、コンドルラプトル、マーショサウルス)が提唱されている。確かに前肢はやや長めの気がする。このキャストは新しいバージョンだろうか。

メインディッシュの1つ、ネオヴェナトル。躍動的な良いポーズだが、一枚の写真に収めるにはどのアングルが良いか悩ましい。ネオヴェナトルは決して新しい恐竜ではなく、ずっと前から知られているが、イギリス国外に出たことがないために日本では一般に馴染みが薄いだろう。NHKの「イギリス恐竜図鑑」で初めて見た人もいるかもしれない。今回、ネオヴェナトル科を紹介するのが趣旨ではないとしても、メガラプトルとともに展示された意義は大きく、感慨深い。
ワイト島の復元骨格をスキャンしたデータを送信し、日本で3Dプリンターで製作したという所は、時代を感じさせる。ということは、ワイト島の復元骨格をそっくり移設したわけだから、昔の復元のままということになる。
ネオヴェナトルは最初、アロサウルス科と考えられたので、欠けている部分はアロサウルスを参考にしているはずである。その後の研究の進展により、アロサウルス科→最も基盤的なカルカロドントサウルス類→最も基盤的なネオヴェナトル科とされてきた(過去の記事「ネオヴェナトル」)。

ネオヴェナトルの頭骨で見つかっているのは前方1/3 つまり吻の部分であり、眼窩や後頭部は見つかっていないので、アロサウルスそっくりに復元されている。涙骨の角状突起がここまで似ているかどうかはわからない。また眼窩の周り、つまり「目つき」もアロサウルスそっくりになっている。
少し下からあおる角度で見えているが、アロサウルスと同じように眼窩の上がオープン、つまり涙骨と後眼窩骨が離れているように作られている。進化したカルカロドントサウルス類では涙骨と後眼窩骨が閉じて、supraorbital shelf (眼窩の上の眉のような部分)を形成する。つまりギガノトサウルスやカルカロドントサウルスのような「目つき」になるわけである。白亜紀前期のコンカヴェナトルでも、後眼窩骨がひさし状に発達して、すでにカルカロドントサウルス類の「目つき」をしていることを考えると、ネオヴェナトルの段階でも、実は後眼窩骨がある程度発達していたかもしれない。ネオヴェナトルは少なくともカルカロドントサウリアであり、アロサウルスよりはカルカロドントサウルス類に近いわけである。

頭骨を傾けたポーズなので、真横から撮るには右側からか。前上顎骨の歯が5本なのはアロサウルスとネオヴェナトルだけで、最も近縁と考えられた。これは現在の系統関係では収斂とされる。それにしてもすばらしい顔ですね。

最近の知見を反映したらしいメガラプトル。今ひとつ狙いがわからない肉食恐竜とみえる。
つづく
コメント ( 2 ) | Trackback ( 0 )
メトリアカントサウルスはシンラプトルと似ているか
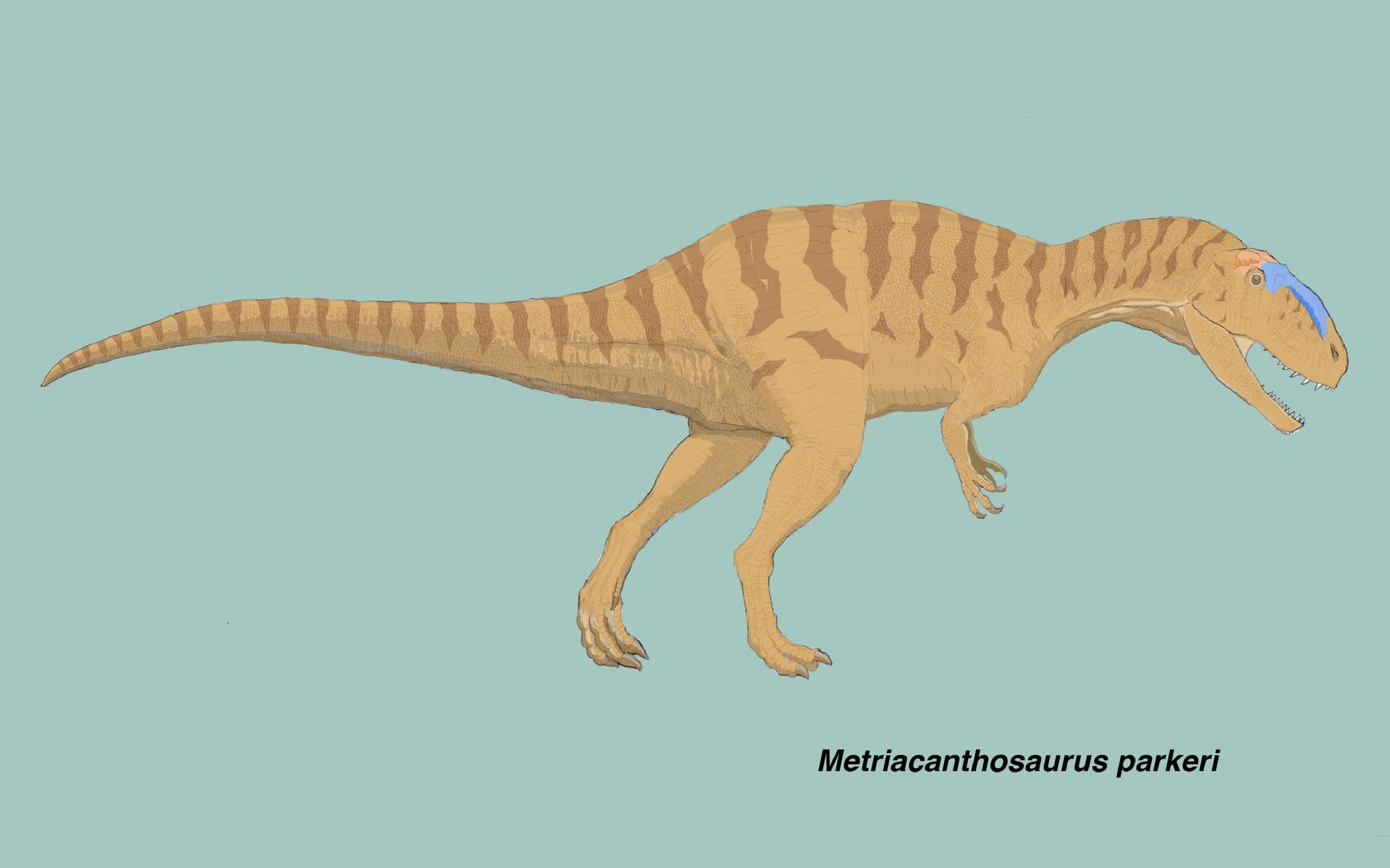
大きい画像
これまで見てきたように、シンラプトルとヤンチュアノサウルスの違いは、腰帯の形質によく表れている。ヤンチュアノサウルスの共有派生形質4つのうち2つは腰帯の形質であり、メトリアカントサウルス亜科の共有派生形質6つのうち3つが腰帯の形質である。そのため、メトリアカントサウルス科の中の分類は、ある程度は、腰帯の形質で読み解くことができる。
それでは、メトリアカントサウルスはどうなのだろうか。メトリアカントサウルスは、常にシンラプトルと共にメトリアカントサウルス亜科に含まれる。腰帯もさぞかしシンラプトルと似ているのだろう、と思って形質データを調べてみた。
メトリアカントサウルスは、胴椎、尾椎、腰帯、大腿骨、部分的な脛骨しか見つかっていない。しかも実は、腰帯の保存状態はよろしくない。断片的な腸骨、恥骨、座骨があるだけである。それでもメトリアカントサウルス科の共有派生形質2つは確かに保持している。298(1) 座骨の断面がハート形(後方にフランジ)と300(1) 座骨の遠位端が癒合している、である。
しかし、メトリアカントサウルス亜科の共有派生形質3つのうち、280(腸骨の後寛骨臼突起の後縁の形)と285(恥骨ブーツの角度)はその部分が保存されておらず、「?」となっている。293(1)(座骨の軸が腹側に曲がっている)は満たしていることになっているが、メトリアカントサウルスの座骨は途中で割れており、遠位の部分はわりと直線的にもみえる。
さらに、ヤンチュアノ側の共有派生形質である281( 恥骨の閉鎖孔などのpuboischiadic plateの状態)は保存されておらず「?」であり、299(座骨の遠位端の形)も「?」となっている。この座骨の遠位端の形は、Walker (1964) の図ではヤンチュアノサウルスのように三角形ではなくシンラプトルと似ているようにみえる。しかしスコアは「?」になっている。
つまりメトリアカントサウルスの腰帯の形質は「?」が多く、メトリアカントサウルス科に含まれるのはいいとして、その中でヤンチュアノサウルスよりもシンラプトルと似ているとは言い切れないようだ。メトリアカントサウルス亜科の共有派生形質6つのうち、腰帯以外の3つは上顎骨、胴椎、脛骨である。メトリアカントサウルスは184(1)(前方の胴椎のキール)と323(1)(脛骨のfibular crestが膨らんでいる)には該当する。総合的にはメトリアカントサウルス亜科に含まれるが、腰帯だけを見ても今ひとつ確認できないということになる。腰帯の図解をしようとしたのに残念である。
当然ながらこの「?」の多さを見ると、メトリアカントサウルスを模式種にしても、他の標本と十分比較できないと感じる。やはり、シンラプトルやヤンチュアノサウルスの仲間という方がわかりやすい。
参考文献
Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012). The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300. Supplementary material
Weishampel, D. B., Dodson, P. & Osm´olska, H. (1990). The Dinosauria. University of California Press, Berkeley, 733 pp.
コメント ( 0 ) | Trackback ( 0 )
恐竜倶楽部30周年記念事業

歴史ある恐竜倶楽部の30周年記念事業が、無事終了しました。講演会の演者の皆様、運営スタッフの皆様、会場の方々、参加者の方々、皆さんありがとうございました。今回も運営のお手伝いをさせていただきました。
また30周年の記念品トートバッグのイラストに、他の2名の方と共に、シンラプトルを採用していただきました。大変光栄に思います。全体として実用的なデザインになったと思います。
展示販売コーナーでは、担当の今富様をはじめ貴重な機会を与えていただき、ありがとうございました。また、シンラプトルその他のイラストをお買い上げいただいた皆さま、ありがとうございました。
コメント ( 0 ) | Trackback ( 0 )