獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2016年5月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | 31 | ||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク





| URLをメールで送信する | |
| (for PC & MOBILE) | |
リスペクトの法則
「イギリス恐竜図鑑」を録画したものを見返したところ、メガロサウルスの歯骨、上顎骨、前上顎骨を並べた後に、頭骨の復元図が描かれていた。画面のキャプチャーは載せられないが、これはどうも吻の形や鼻孔が大きい点など、ネオヴェナトルの頭骨に似ている。メガロサウルスの正確な頭骨図というものはないので、ネオヴェナトルで代用し、ネオヴェナトルの前上顎骨をもってきたのだろうか。(録画された方は確認してみていただきたい。「実は私はMaverick TVの製作関係者で事情を知っているが、こういうことなんですよ」という方はご一報下さい。)
確かに、ネオヴェナトルの吻の先端は斜めにとがっている(前上顎骨の前縁が傾いている)ので、その点でメガロサウルス類と似ている。これでいいだろう、ということか。ただしこの頭骨復元図は、番組中のメガロサウルスのCGとは異なる。CGのメガロサウルスはもう少しティラノ的というか、吻の先端が垂直に近い角度だった。考えすぎかもしれないが、これも、実は意味があると思われる。メガロサウルスの前上顎骨は見つかっていないので、吻の先端の形は実際にはわからない。しかし、メガロサウルスでは他のメガロサウルス類とは異なり、上顎骨の前方突起が短いことから、吻の先端は他のメガロサウルス類ほど斜めではなく、垂直に近い可能性がある。そのことについては論文にはないが、Wikipediaをよく読むと書いてある(もちろん執筆者は不詳)。よってメガロサウルスの顔は、トルボサウルスやドゥブレウイロサウルスとは異なってもよいわけである。
そこで昔考えたことのある、「獣脚類リスペクトの法則」を思い出したので、雑文として書いておく。ある地域では、なぜか時代が異なっても同じような特徴をもった恐竜が現れるという法則である。
北アメリカでは、ジュラ紀後期の覇者アロサウルスをリスペクトするかのように、白亜紀後期のゴルゴサウルスやダスプレトサウルスなどが涙骨の角状突起を発達させ、
新疆ウイグルでは、ジュラ紀中期のモノロフォサウルスをリスペクトするかのように、全く別系統のティラノサウロイドであるグァンロンが正中のとさかを発達させ、
南米などのゴンドワナ地域では、白亜紀中頃に衰退したカルカロドントサウルス類をリスペクトするかのように、アベリサウルス類がごつごつした顔面や頭蓋天井を発達させ、(眼窩下突起もあるし)
イギリスでは、ジュラ紀中期のエウストレプトスポンディルスなどのメガロサウルス類をリスペクトするかのように、白亜紀前期のネオヴェナトルが斜めにとがった吻部を発達させ、(歯骨の3番目の歯槽まで似ている)
という具合である。もちろん実際には、これは様々な時代に様々な系統で収斂進化が起きているということだろう。たとえば肉食恐竜がその行動様式や生態上、邪魔にならない部分(涙骨や鼻骨)に装飾構造を発達させることが可能、ということだろう。捕食者として強く咬むためには頭骨の丈が高くなり、角質で強化するための表面構造も発達するだろう。スピノサウルス類で前上顎骨と上顎骨の間がくびれているのは、コエロフィソイドの形質を受け継いだのか独自に獲得したのか、わからないようである。独立して獲得したのだとすれば発生プログラムの都合ではないだろうか。
コメント ( 0 ) | Trackback ( 0 )
ドゥリアヴェナトル
ドゥリアヴェナトルは、ジュラ紀中期バジョシアン(Inferior Oolite)にイギリスのドーセット州に生息したメガロサウルス類で、2008年に新属として記載された。
ドーセット州のInferior Ooliteから発見された獣脚類の頭骨の一部は、Owen (1883) によってメガロサウルス・バックランディMegalosaurus bucklandi (iが1つ)として記載されていた。しかしWalker (1964)は、この標本はメガロサウルスとは歯の数が異なることに気づいた。そこでWaldman (1974) は、メガロサウルス属の新種メガロサウルス・ヘスペリスMegalosaurus hesperis とした。その後の研究者はこれを疑問とし、属名をカッコでくくった‘Megalosaurus’ hesperisと記してきたが、詳細な研究がなされていなかったので、メガロサウルスとの正確な比較はできず、特徴も定まっていなかった。Benson (2008)はこの標本を詳細に記載し、メガロサウルスとは異なる形質をもつことから、新属ドゥリアヴェナトルを設けてドゥリアヴェナトル・ヘスペリスDuriavenator hesperisとした。ドゥリアはドーセットの古名で、「ドーセットの狩人」である。
ホロタイプNHM R332は、左右の前上顎骨、右の上顎骨、鋤骨、左右の歯骨、右の上角骨、同定できない骨の断片、分離した歯からなる。上顎骨、歯骨には多くの歯が保存されており、かなり迫力のある化石である。
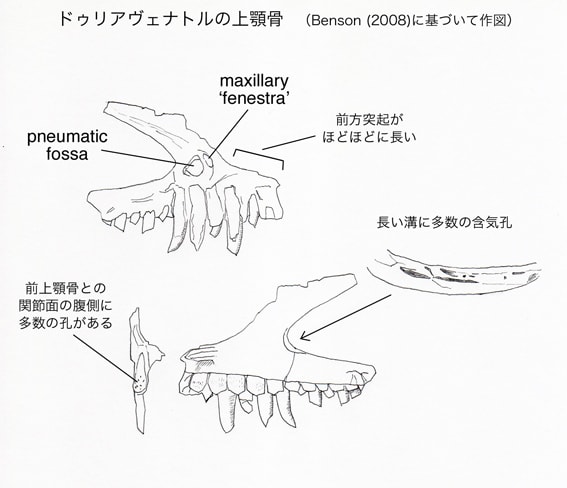
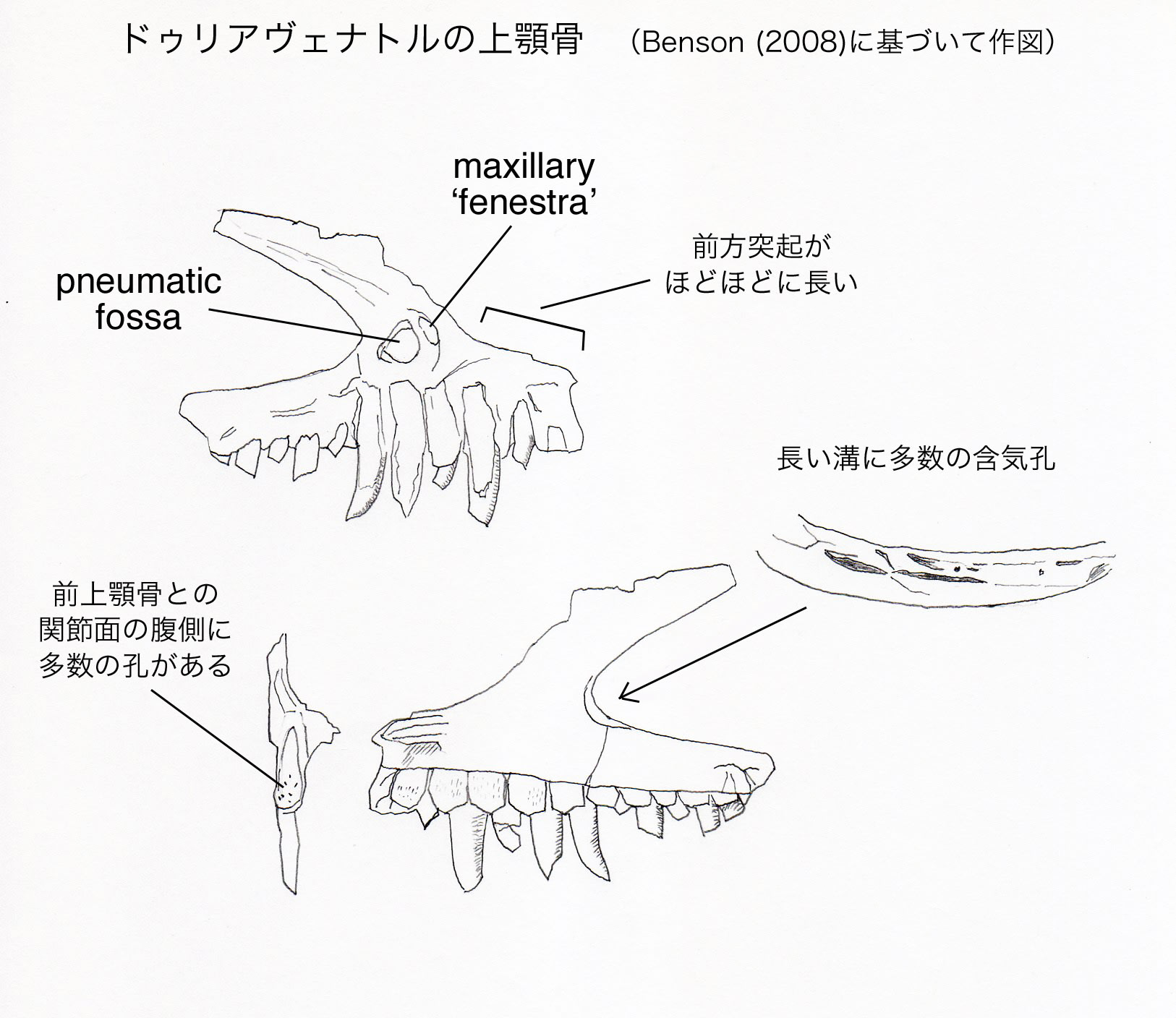
ドゥリアヴェナトルは、上顎骨に以下の固有形質をもつメガロサウルス類である。頬骨突起の背側面に深い溝があり、その中に多数の含気孔がある;前上顎骨との関節面の腹側部分に、多数の小さい孔がある。
前上顎骨
左の前上顎骨の前方部分と右の前上顎骨の後背方部分だけが保存されている。上顎骨が埋まっていたブロックの観察から、Walker (1964)は5本の前上顎歯があったかもしれないとコメントしている。 Waldman (1974)は、2本の歯冠、1本の断面、1本の歯の印象をあわせて4本の前上顎歯をみとめている。これらの観察は、現在では(Waldmanがクリーニングした後なので)確認できない。もしドゥリアヴェナトルが5本の前上顎歯をもっていたとすれば、メガロサウロイドとしてはユニークな特徴となる。ドゥブレウイロサウルスとエウストレプトスポンディルスでは4本であり、スピノサウルス類では6-7本であるという。
前上顎骨の前縁は、腹側縁に対して約50°の角度で後背方に傾いている。この角度はトルボサウルスと近い。ドゥブレウイロサウルスでは約45°、エウストレプトスポンディルスでは約40°であるという。前上顎骨の外側面は保存がよくないが、前方に多数の栄養孔がみられる。
大きい画像
上顎骨
上顎骨の前上顎骨との関節面の腹側に、直径約2 mmの11個の孔がある。これらの孔は、バリオニクスやメガロサウルスなど多くの基盤的テタヌラ類には存在しない。カルカロドントサウルスには孔があるが、孔の分布がドゥリアヴェナトルよりもずっと背方に広がっている。ドゥリアヴェナトルの状態は固有形質と考えられる。
上顎骨の前方突起は、前後の長さと背腹の高さが同じくらいである。これは派生的なカルカロドントサウルス類やシンラプトル類よりも長い。長い前方突起はメガロサウルス上科の共有派生形質とされたことがあるが、アロサウルス、グァンロン、モノロフォサウルス、ネオヴェナトルにもみられる。前方突起から前方に、つまみ状のフランジtab-like flangeが突出している。このフランジは前上顎骨の外側面と重なっていたと思われ、バリオニクスにおいて記載されているanterior pegと相同なものかもしれないという。同じような構造はトルボサウルスにみられ、ドゥブレウイロサウルスにはみられない。
上顎骨の外側面は剥がれて、反対側(カウンターパート)の石板に付着している。そのため外側面の表面構造や孔については観察できない。前眼窩窩の前端部分も骨が剥がれていて、2つの含気性の腔所が見えている。このうち前方のものがmaxillary 'fenestra'であり、貫通したfenestraではなく内側が閉じた凹みとなっている。後方の穴は(maxillary fenestraのように見えるが)頬骨突起の背側面にある凹みの延長であるという。この含気性の凹みは、縦に長い溝の形をとり、その中に多数の孔がある。この位置の含気性の凹みは、他の基盤的テタヌラ類にもみられるが、それらの種では卵形で長く延びていない。そのため、ドゥリアヴェナトルの状態は固有形質と考えられる。
上顎骨の11個の歯槽のすべてに、成長中あるいは成長した歯が保存されている。Walker (1964) は全部で15-18本の歯があったと推定した。ドゥリアヴェナトルの上顎骨はドゥブレウイロサウルスと最も似ている。ドゥブレウイロサウルスとの比較からは、14-16本と推定される。多くの獣脚類と同様に、歯冠は後方に反っており、鋸歯がある。弱いエナメルのしわがあり、前縁と後縁の間にバンド状に延びている。歯間板は五角形で癒合していない。歯間板は上顎骨の外側壁と同じレベルまで伸びている。歯間板の表面にはわずかに孔や条線がある。
大きい画像
歯骨
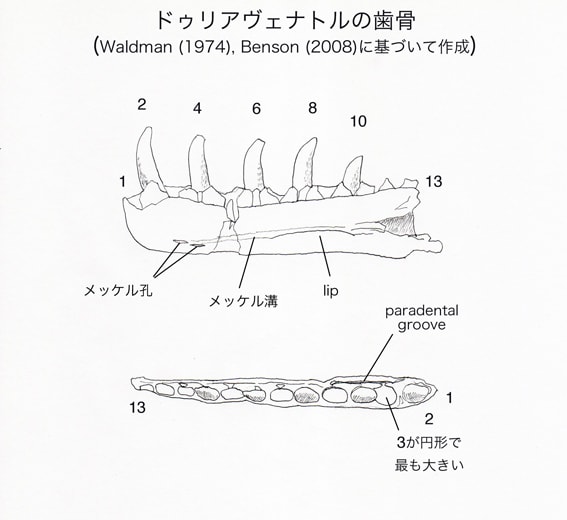
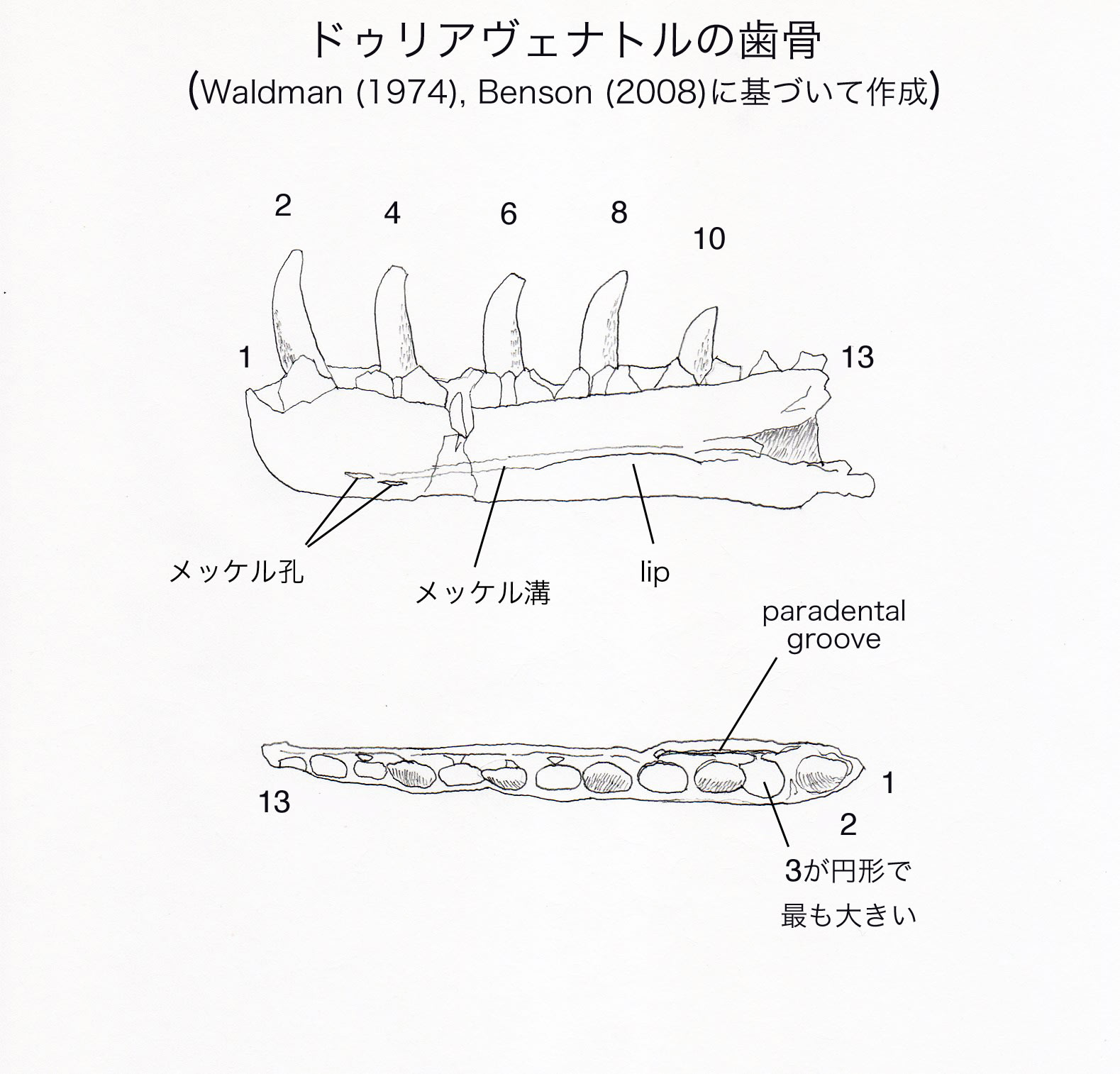
左右の歯骨があるが、右の歯骨の方がより完全で、最初の13個の歯槽が保存されている。Waldman (1974)は全部で17-18本と推定したが、Benson (2008)ではアロサウルスとの比較から14-15本と推定している。右の歯骨では、2、4、6、8、10番目の歯槽に成長した歯がある。4、5、6、8番目の歯槽の内側の歯間板の間に交代歯がある。
最初の3個の歯槽は円形に近く(subcircular)、3番目がすべての歯槽の中で最も大きい。4番目以後の歯槽は長方形に近く(subrectangular)、後方へいくにつれて小さくなっている。拡大した歯槽を収めるため、歯骨が3番目の歯槽の位置で側方に膨らんでいる。これはマグノサウルスなど、メガロサウルス以外のメガロサウルス類と同様である。
右の歯骨の2番目の歯は、メガロサウルス類の歯骨の前方の歯としては珍しく保存されたものである。これは後方の歯よりも長く、断面が円形で、前背方に傾いている。より後方の歯では前方の稜縁が歯冠の高さの1/3くらい延びているが、2番目の歯では短い距離しか延びていない。歯骨の最も前方の歯は、コエロフィソイドでも似た形態を示す。また同じように拡大した、円形に近い歯槽をもつ他の獣脚類、例えば他のメガロサウロイド、ディロフォサウルス、アクロカントサウルス、ネオヴェナトルでも似た形態かもしれないという。この傾向が極端に発達した例がマシアカサウルスにみられる。
歯列の内側には五角形の歯間板がある。歯間板の表面は弱く孔のある構造をしている。歯間板と歯骨の内側壁の間にparadental groove がある。paradental grooveは前方では広く、後方で狭くなっている。
メッケル溝の腹側縁は背側縁よりもはっきりしていて、突出した唇状部lipをなしている。メッケル溝の前端付近に2個のメッケル孔がある。2個とも前後にのびた楕円形をしており、1個がもう1個の前方でわずかに背方に位置している。
他のメガロサウルス類との違い
ドゥリアヴェナトルは、メガロサウルスの歯骨とは異なるいくつかの特徴を示す:拡大した円形に近い3番目の歯槽;円筒形に近い(subcylindrical)、前方に傾いた前方の歯骨歯;前方のメッケル孔が後方のメッケル孔のほとんど前方に位置する;歯間板の丈がより低い;メッケル溝の腹側に顕著な唇状部lipがある。
ドゥリアヴェナトルは、同じドーセットのバジョシアン前期のマグノサウルスとは、歯骨だけが比較できる。ドゥリアヴェナトルの歯骨はマグノサウルスの歯骨とは、側方から見て前端がそれほど強く拡大していないこと、外側面の縦の溝longitudinal grooveがそれほど深くないこと、メッケル溝の腹側に顕著な唇状部があること、で異なるという。
マグノサウルスは後肢の長さ1315 mmと比較的小型であるが、成体であるという。一方ドゥリアヴェナトルは、メガロサウルスに匹敵するくらいの大きさと考えられている。
メガロサウルスは「3番目の歯槽が拡大していない」というので、それじゃ他のメガロサウルス類はどうなのか、と思って調べているわけである。確かに3番目の歯槽が円形で大きいのはわかる。しかし破損や変形もあることを考えると、微妙といえば微妙な気もする。paradental grooveは、ドゥリアヴェナトルの場合、確かにわかる。
細かいことをいえば、上顎骨の歯の数はドゥブレウイロサウルスと比較しているのに、歯骨の歯の数はアロサウルスと比較して推定しているが、これはどうなのか。しかもメガロサウルスの論文では、歯骨の歯の数はドゥブレウイロサウルスと比較している。また2番目の歯が後方の歯よりも長いというが、4番目、6番目の歯は明らかに先端が欠けている。先端までいれるとそれほど差はないようにみえる。わずかに長いのかもしれないが微妙である。
メガロサウルスとドゥリアヴェナトルは、時代も異なり、歯の数以外にも3番目の歯槽や、前方突起の長さなどいろいろ違うことがわかる。しかしドゥリアヴェナトルとマグノサウルスは時代も場所も近く、1個体同士の比較であることを考えると、その違いは結構微妙なようにも思える。
参考文献
Benson RBJ. (2008) A redescription of 'Megalosaurus' hesperis (Dinosauria, Theropoda) from the Inferior Oolite (Bajocian, Middle Jurassic) of Dorset, United Kingdom. Zootaxa 1931: 57-67.
Waldman, M. (1974) Megalosaurids from the Bajocian (Middle Jurassic) of Dorset. Palaeontology, 17, 325-340.
コメント ( 0 ) | Trackback ( 0 )
100円ラプトル

ダイソーなどの100円ショップで売っている商品で、他に100円ティラノや100円ケラトもある。いくら私でも100円のおもちゃに突っ込みを入れる気はないが、ちょっと面白いので。。。
種名がなく、ヴェロキラプトルなのかデイノニクスなのかもよくわからない。それより手に注目していただきたい。足の第2指の他に、手の第1指も持ち上げているのが特徴である。これを作った人は何か勘違いをしているのではないか?手の指を反らせてどうする。なにか印を結んでいるのか、東南アジアの民族舞踊か。。
コメント ( 0 ) | Trackback ( 0 )
黒須さんのサイエンスカフェ
5/8の午後2時から高田馬場の10°cafe で開催された、黒須さんのクラウドファンディングのサイエンスカフェに参加しました。一言で言うとGW連休中で一番充実した一日となりました。
中学生の時の内モンゴル恐竜ツアーの話から、中国に留学した経緯、中国地質大学の博物館や学生食堂、大学生活の様子などが紹介されました。最近の恐竜研究の例として、成長曲線などのボーンヒストロジー、アンキオルニスの体色の復元、トリケラトプスの復元姿勢などの機能形態学を紹介した後、ご自身の研究テーマの解説となりました。中国では恐竜化石の多さに対して研究者の数が少なく、次から次へと記載して行かなければならない状況ということです。
甘粛省のゆ(輸のつくり)井子盆地から発見されたドロマエオサウルス類は、頭骨では前頭骨のみ、胴椎4個、仙椎、腰帯、尾椎が少し、肩甲骨や前肢の一部、後肢の一部があるそうです。
前頭骨については、Currie (1987) や最近のボレオニクスの論文の図と比較しながら、ボレオニクスよりもさらに細長いということでした。上側頭稜supratemporal ridgeは恐らく摩耗のため、薄くてほとんど見えない。前方の鼻骨との関節面はわりと狭い。ヴェロキラプトルのようにわりと細長い頭骨だったらしいということです。さらに長い頭骨としてブイトレラプトルの写真もありました。
各地の地質公園の紹介として、まず山東省の諸城のボーンベッドを建物で覆った様子と大量の化石群。特に「諸城ティラノサウルス館」には、明らかにティラノサウルス類とわかる骨があちこちに転がっており、まだ手つかずの状態だそうです。諸城にはズケンティランヌスの他に、あと2種くらいはティラノサウルス類があるようです。ズケンゴサウルス、ズケンケラトプスなども出ました。
遼寧省の熱河生物群の紹介と、朝陽市鳥化石国家地質公園。ここには鳥の化石だけで200-300もあり、とても全部は撮影できないそうです。ドロマエオサウルス類の化石もいくつか紹介されました。マンチュロケリスなどのカメ、孔子鳥などの標本。
あと最近の岩手県久慈層群での獣脚類の歯の発見に立ち会った話。2センチ弱で鋸歯がないという歯ですね。
予定よりも時間が押したため、リターンの受け取りの後は場所を変えて、駅前で2次会となりました。研究生活では、飲み会が多すぎるのが現在の悩みだそうです。
その後さらに、私は前頭骨の関節面の写真や、安徽省の博物館にあるドロマエオサウルス類の全身化石の写真などを見せていただき、またいろいろなお話を伺うことができました。黒須さんはもとより参加者の皆さん、どうもありがとうございました。
コメント ( 2 ) | Trackback ( 0 )