獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2018年4月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | |||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ヤンチュアノサウルス・ジゴンゲンシス(スゼチュアノサウルス・ジゴンゲンシス)
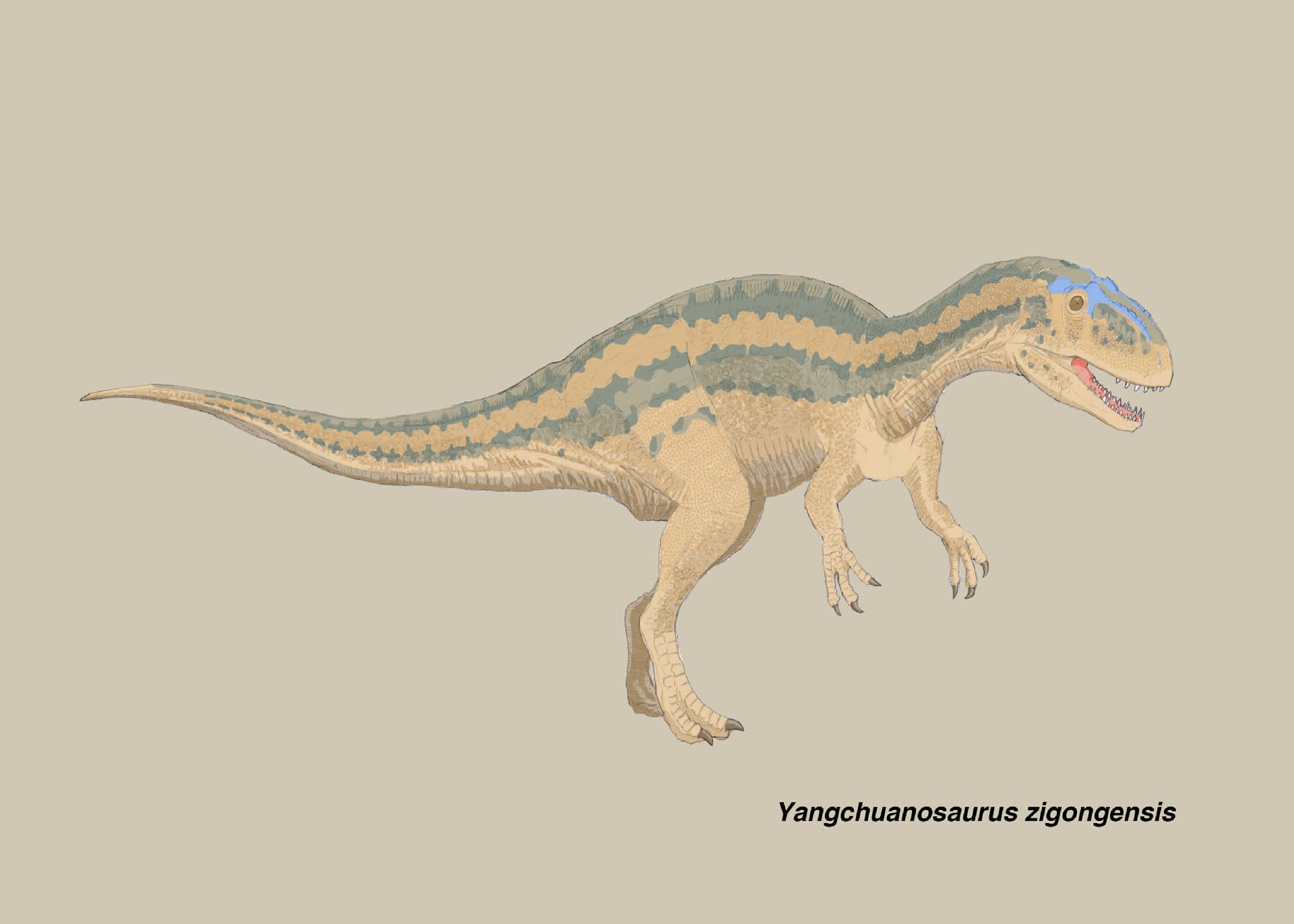
大きい画像
ヤンチュアノサウルスやシンラプトルに比べて、具体的なイメージがつかめないのがスゼチュアノサウルスである。中国の恐竜展などで復元骨格を見るたびに、どこを観察すればいいのか戸惑ってしまう。しかもある意味残念なことに、Carrano et al. (2012) の系統研究で、この「四川竜」の属名は失われてしまった。
スゼチュアノサウルスの歴史
歴史的経緯については、Wikipedia(英語)等にも書いてあるので簡潔にメモする。
Young (1942)が記載したスゼチュアノサウルス・キャンピの模式標本は、4つの分離した歯化石で、同定できる特徴がないために、現在は疑問名とされている。そのため新たに発見された化石にスゼチュアノサウルスの属名を用いることは不適切である。
四川省自貢のジュラ紀後期の地層から発見された部分骨格CV00214は、1978年にスゼチュアノサウルス・ヤンドネンシスとされ、1983年にスゼチュアノサウルス・キャンピとされたが、上述の理由で長年疑問視されていた。頭骨はなく、歯は含まれていない。
さらにGao (1993)は、四川省自貢のジュラ紀中期の地層から発見された部分骨格に、スゼチュアノサウルス・ジゴンゲンシスと命名した。Chure (2001) は、CV00214とスゼチュアノサウルス・ジゴンゲンシスを同一種として、(スゼチュアノサウルスは使えないので)新しく命名すべきであると考えた。
Carrano et al. (2012) の系統解析の結果、CV00214はヤンチュアノサウルス・シャンヨウエンシスと姉妹群となり、それらとスゼチュアノサウルス・ジゴンゲンシスが姉妹群となった。つまり系統関係上も“スゼチュアノサウルス”は解体された。この結果を受けてCarrano et al. (2012)は、同じ地層から出ていることからもCV00214はヤンチュアノサウルス・シャンヨウエンシスと同一とし、より古い地層から出ているスゼチュアノサウルス・ジゴンゲンシスは別種ヤンチュアノサウルス・ジゴンゲンシスとした。新しい属名が必要になったがヤンチュアノサウルス属の別種とすることで系統関係の近さを表したといっている。
ここで気になる点が2つある。1つは、種レベルの違いか属レベルの違いかの判断はやはり、研究者によって異なることである。スゼチュアノサウルスの代わりに、新しい属名がついた方がアマチュア的には楽しかった。
もう1つは、中国四川省は獣脚類化石の宝庫であるが、あまり欧米の研究者がアクセスできていないようにみえることである。Carrano et al. (2012) は、系統解析に用いた標本の90%は、3人の共著者の誰かが直接観察していると述べている。First handで見た種類は太字で表記している。しかし四川省のヤンチュアノサウルスやスゼチュアノサウルスの標本は、直接見ておらず、文献の記述と写真・図版からデータをとっているようである。四川省の研究者が認めているかどうか知らないが、これでは納得しないかもしれない。本当は、中国側と欧米側が共同研究して、結論を出してほしいところである。
ジゴンゲンシスの特徴
ヤンチュアノサウルス・ジゴンゲンシス(スゼチュアノサウルス・ジゴンゲンシス)は、中国四川省自貢市大山舗のジュラ紀中期の地層から発見されたメトリアカントサウルス類で、Gao (1993)によって記載された。自貢恐竜博物館の建設工事中に発見された大量の恐竜化石の1つで、推定6mの肉食恐竜である。
ホロタイプZDM9011は胴体のかなりの部分を含む部分骨格で、連続した完全な10個の頸椎、13個の胴椎、5個の仙椎、不連続な25個の尾椎、左の上腕骨、尺骨、橈骨、橈側骨、中手骨、指骨、右の肩甲骨、完全な腰帯からなる。その他に同時に発見された参照標本としてZDM9012:左上顎骨、ZDM9013:10本の分離した歯、ZDM9014:右の大腿骨、脛骨、腓骨がある。
Gao (1993)はジゴンゲンシスの特徴として、前部・中央部の頸椎は後凹型、後部の頸椎は平凹型で腹側にキールがある、後部尾椎の前関節突起が長い、上腕骨の三角筋稜がよく発達している、橈骨/上腕骨の比率が56%、第IV中手骨が残存している、腸骨は特に丈が低い、恥骨ブーツが小さい、座骨の遠位端が拡がっている、などをあげている。(中国語と英語で内容が微妙に異なる。)
一方Carrano et al. (2012)は、頸椎の特徴だけをあげている。Holtz et al. (2004) に従い、前方の頸椎2-4 だけが後凹型、残りは両平面型であるテタヌラ類としている。後方の頸椎の関節面が平面的ということだろう。Gao (1993)のあげた特徴のいくつかは、メトリアカントサウルス科やヤンチュアノサウルスの特徴に含まれた。
Gao (1993)は、スゼチュアノサウルス・キャンピとされたCV00214とジゴンゲンシスの違いについて記している。ジゴンゲンシスでは、後部頸椎が平凹型で腹側にキールがある;三角筋稜が特に発達している;第IV中手骨が存在する。一方CV00214は、頸椎が後凹型でキールがない;三角筋稜があまり発達していない;第IV中手骨がない。
ヤンチュアノサウルス・シャンヨウエンシスのホロタイプでは前肢は保存されていないが、CV00214がこれと同一種とすれば、完全に3本指だったことになる。
なぜスゼチュアノサウルスがヤンチュアノサウルスになったのかについては、次の記事に持ち越すことにする。
参考文献
Gao, Y. (1993). [A new species of Szechuanosaurus from the Middle Jurassic of Dashanpu, Zigong, Sichuan]. Vertebrata PalAsiatica, 31, 308-314 [In Chinese].
Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012). The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300.
Shangshaximiao Formation Xiashaximiao Formation
コメント ( 0 ) | Trackback ( 0 )
浅草でメガロサウルス教室

責任上、メガロサウルス類教室も行かないわけにいかないので、行ってきた。
楽しかったです。
トルボサウルスは、この時代の凄まじい動物の一つですね。
私の好きなドゥブレウイロサウルスも登場したし。懐かしい。私が昔、ドゥブレウイロサウルスの記事を書いた時には、日本語で表記した文章自体がほとんどなかったので、ローマ字読みしておいた。ダブルーオとか、デュブルイヨなど色々な表記があるが、フランス語をカタカナで表記すること自体に無理がある。好きに読めば良いのです。この恐竜は今でも知名度の低さはほとんど変わっていないが、ヨーロッパの恐竜本(Darren Naishなど)には結構登場している。
復元教室は1日楽しすぎて、月曜からが憂鬱になるのが難点です。
コメント ( 0 ) | Trackback ( 0 )
「羽毛恐竜 恐竜から鳥への進化」

大島英太郎作・真鍋真監修ということで、前作「とりになったきょうりゅうのはなし」の続編でしょうか。
楽しい絵本です。表紙をめくると、とびらの、科博でデイノニクスを見学する親子の絵が秀逸で、ぐっと引き込まれます。内容はシノサウロプテリクスの発見から、シノルニトサウルス、ミクロラプトル、クリンダドロメウスやユーティラヌスなど様々な羽毛恐竜、ディスプレイやシチパチの抱卵、鳥への道筋が単純ではないこと、翼竜や被子植物の繁栄との関係などにわたっています。
恐竜絶滅のページをめくると、パッと明るい色調で現生鳥類の繁栄が描かれていて印象的です。最後はツバメの子育てを見上げる子供の絵で、構成としてオープニングとラストの絵がよく考えられていると思いました。
真鍋先生の解説だけでなく、大島英太郎氏による解説ページがあるのが良いですね。楽しく読めました。
コメント ( 0 ) | Trackback ( 0 )
シンラプトル2018 (2)
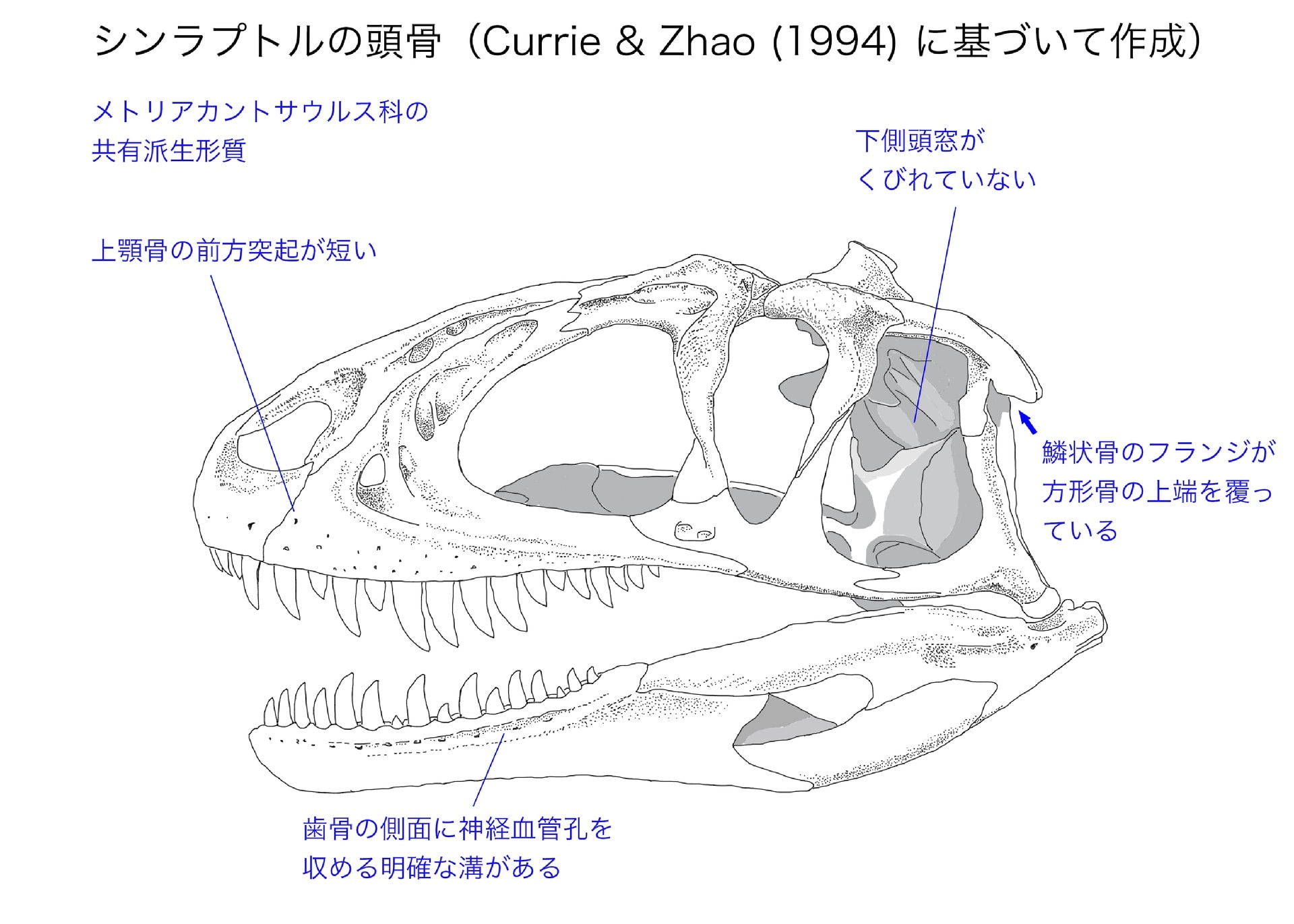
メトリアカントサウルス科の共有派生形質
メトリアカントサウルス科は12の明確な共有派生形質でまとめられるが、細かい形質が多く難解なものもある。その中でわかりやすいのは、前述した第IV中手骨である。「第IV中手骨が存在し、第IV指の指骨と第V指が失われている」に該当する。アロサウルス上科の中では主にメトリアカントサウルス科だけにみられる形質で、シンラプトル、ヤンチュアノサウルス・ジゴンゲンシス、シュワンハノサウルスで確認される。

また少し難しそうだが重要な形質として、軸椎の間椎心intercentrumの向きがある。(頸椎のうち第1頸椎を環椎、第2頸椎を軸椎という。それぞれ本来は間椎心と椎心(椎体)からなっていて…という環椎・軸椎の構造については、人体解剖学や比較解剖学の本に載っている。)
シンラプトルでは歯状突起odontoidは分離して見つかっている。軸椎の腹側では間椎心と椎心(椎体)が結合している。多くの獣脚類(ディロフォサウルス、ケラトサウルス、カルノタウルス、ピアトニツキサウルス、アロサウルス)では軸椎の間椎心の腹側面が水平であるが、シンラプトルでは前背方に傾いている。Currie & Zhao (1994) は、ジュラ紀の獣脚類では他にヤンチュアノサウルスやモノロフォサウルスなど2,3の種にしかみられないと述べている。Carrano et al. (2012) によると、この形質はマーショサウルスを含むいくつかの基盤的なテタヌラ類にみられるという。いずれにしてもメトリアカントサウルス科は「軸椎の間椎心の腹側面が前背方に傾いている」を共有する。
この軸椎の形態は、頸椎を後頭顆の下に寄せることで頭骨をよく支える役割があると考えられている。同じような適応はティラノサウルスやタルボサウルスにもみられるといっている。つまりこれは原始的な形質ではなく、大きな頭骨を支えるための先進的な形質ともいえるのではないか。シンラプトルも頑張っている。

メトリアカントサウルスやシャモティラヌスが近縁とされたということは、腰帯の形質が効いているにちがいない。シャモティラヌスはどこがシンラプトルと似ているのか。
メトリアカントサウルス科の共有派生形質の一つは座骨の形態で、「左右一対の座骨軸の断面がハート形」、つまり正中部分がフランジ状に後方に突出していることである。Currie & Zhao (1994) は座骨の後背方にcrestがあるといっていて、同じくらい発達しているのはヤンチュアノサウルスだけであると述べている。もう一つは「座骨の遠位端が癒合している」ことである。これはシャモティラヌスでは確認できない。
また腸骨の後寛骨臼突起の後縁が直線的であることと、恥骨ブーツが強く後背方に傾いている(恥骨軸と恥骨ブーツの角度が60度より小さい)ことは、メトリアカントサウルス亜科の共有派生形質となっている。
メトリアカントサウルス、シンラプトル、ヤンチュアノサウルスはいずれも胴椎の神経棘が中程度に長いが、そのことはメトリアカントサウルス科の確実な共有派生形質には含まれていない。(ACCTRANという方式の場合のみ入る。)メトリアカントサウルス科の中にはヤンチュアノサウルス・ジゴンゲンシスのように胴椎の神経棘があまり長くないものもいるので、共通祖先の形質ではなく、一部の種類が獲得した形質ということだろう。これもアクロカントサウルスのようなカルカロドントサウルス類との収斂的な傾向かもしれない。
シンラプトルのホロタイプは推定全長7.2 m であるが、脳函の骨同士、脊椎の神経弓と椎体、個々の仙椎同士などが完全に癒合していないことから、亜成体と考えられている。成体はもう少し大型だったかもしれない。全長7.2 mに対して頭骨は90 cm と頭がかなり大きい。中足骨はアロサウルスよりも長い。ただし亜成体であることを考えると、成体はアロサウルスと同じくらいの可能性があるという。
コメント ( 0 ) | Trackback ( 0 )
シンラプトル2018(1)

大きい画像
シンラプトル科からメトリアカントサウルス科へ
シンラプトルやヤンチュアノサウルスの仲間は伝統的にシンラプトル科Sinraptoridaeと呼ばれてきたが、Carrano et al. (2012) のテタヌラ類の系統解析の結果、歴史的に古いメトリアカントサウルスがシンラプトル科に含まれたため、メトリアカントサウルス科Metriacanthosauridaeと呼ばれるようになった。ただし、新しく含まれたメトリアカントサウルスやシャモティラヌスなどは化石が断片的で、明確なイメージがない。依然としてこのグループの代表は中国のシンラプトルやヤンチュアノサウルスであり、はるかに完全な全身骨格による明確なイメージがある。また何度も日本の恐竜展などで公開されているので、昔からの日本の恐竜ファンにとっては、シンラプトル科の方が馴染みがあるだろう。ただし最近の書籍ではメトリアカントサウルス類になっているはずなので、シンラプトル科を知っているのはオッサンの証拠になるかもしれない。
実際には、「メトリアカントサウルスがシンラプトル科に含まれた」という表現は正確でないかもしれない。分岐分析の結果、昔のシンラプトル科のメンバーを含む、拡張されたクレードが出現したということで、とらえ方が異なるのだろう。
メトリアカントサウルス科には、シュワンハノサウルス、シダイサウルス、ヤンチュアノサウルス・シャンヨウエンシス、“スゼチュアノサウルス・キャンピ”とされたCV00214、“スゼチュアノサウルス”・ジゴンゲンシス、メトリアカントサウルス、シンラプトル・ドンギ、シンラプトル・ヘピンゲンシス、シャモティラヌスが含まれた。ただし断片的なシュワンハノサウルスを除外した方がずっと分岐図が安定し解像度も上がった。分岐図はネット上にある。
メトリアカントサウルス科の中には2つのクレード、ヤンチュアノサウルスの仲間とシンラプトルの仲間が含まれる。ヤンチュアノサウルスの仲間は、“スゼチュアノサウルス”の使用が無効とされ、全てヤンチュアノサウルス属となった。一方、シンラプトルの仲間はメトリアカントサウルス亜科Metriacanthosaurinaeとされ、メトリアカントサウルス、シンラプトル・ドンギ、シンラプトル・ヘピンゲンシス、シャモティラヌスが含まれている。
シンラプトル科は、シンラプトルを詳細に記載したCurrie & Zhao (1994) によって樹立された。そこでは、シンラプトルとヤンチュアノサウルスがシンラプトル科に含まれるとしている。しかしその当時は、多くのカルカロドントサウルス類の標本がほとんど発見されていなかったので、アロサウルスやケラトサウルスなど、限られた種類としか比較されていない。そのため当時のシンラプトル科の特徴は変わってくると思われる。例えばシンラプトル科の特徴として、上顎骨の前眼窩窩に2個以上の副次的な孔があることをあげている。現在では、アクロカントサウルスやエオカルカリアにも副次的な孔があることは知られているが、シンラプトルほど多くはないようにもみえる。この特徴の位置づけはどうなったのだろうか。(メトリアカントサウルス科の特徴ではない。)一方、シンラプトル科の特徴のうち軸椎の間椎心の傾きについては、メトリアカントサウルス科の特徴として受け継がれている。

シンラプトルの特徴
Currie & Zhao (1994)ではシンラプトル属の特徴として、頭骨がヤンチュアノサウルスよりも長く、丈が低いこと、頬骨に大きな含気孔があること、後眼窩骨に粗面が発達していることなどが含まれていた。Carrano et al. (2012) では、これらの一部(以下の1)2)3))をシンラプトル・ドンギの特徴として採用している。
Carrano et al. (2012) によるシンラプトル・ドンギの特徴は、アロサウロイドのうちで1)下側頭窓が大きく、比較的長くまっすぐな後眼窩骨-鱗状骨バー、2)後眼窩骨の後方突起(鱗状骨突起)が非常に短くみえる、3)口蓋骨の内部に深く含気性が発達している、の3つである。後眼窩骨-鱗状骨バーとは、上側頭窓と下側頭窓の間を仕切る部分でCurrie & Zhao (1994)ではintertemporal bar と呼んでいたが、Carrano et al. (2012)ではpostorbital-squamosal bar と言い換えている。またCurrie & Zhao (1994)によると、後眼窩骨の後方突起は非常に短くみえるが、内側から見るともっと後方まで伸びていて、外側が鱗状骨に覆われているのであるという。そのためCarrano et al. (2012)は短いと言い切っているが、ここでは「短くみえる」としておいた。
原始的なアロサウロイド
シンラプトルはよく、「アロサウルスよりも原始的」と書かれる。どこが原始的なのだろうか。
Currie & Zhao (1994)では、シンラプトルがアロサウルス科よりも原始的な点として、下側頭窓が大きいことから始まって、距骨の上行突起が比較的低いことまで、非常に多くの形質をあげている。
おそらく最もわかりやすいのは第IV中手骨が残っていることで、テタヌラ類は3本指であるが第4指の痕跡があるということである。これはシンラプトル科の特徴には含まれていなかったが、メトリアカントサウルス科の特徴として含まれている(後述)。
またアロサウルス上科の系統進化を論じる研究では必ず言及されるのが、歯間板の状態である。シンラプトルでは、ドゥリアヴェナトル、ピアトニツキサウルス、モノロフォサウルスのような基盤的なテタヌラ類と同様に、歯間板が分離しており、癒合していない。アロサウルス上科の中で、アロサウルス以上の種類では歯間板が互いに癒合している。さらに派生的なカルカロドントサウルス類では、歯間板の丈が高くなっている。メガロサウルス類の中でも、トルボサウルスなどでは歯間板が癒合し、強化されている。一方小型のコエルロサウルス類出身のティラノサウルス類では、歯間板は癒合していない。つまりティラノサウルス類では顎や歯自体は非常に頑丈になっているが、歯間板の強化については原始的な状態である。
シンラプトルの肩甲骨は、太いメガロサウルスと細長いアロサウルスの中間的な形で、遠位端がはっきり拡がっていない点でアロサウルスよりも原始的とされている。
シンラプトルでは頸椎と前方の胴椎の含気腔が単純で、多数の骨梁trabeculae で仕切られていないとある。これもアロサウルス上科全体の進化傾向からみると、カルカロドントサウリアでは頸椎と胴椎の含気性が発達していくので、シンラプトルは原始的ということが納得できる。
シンラプトルがカルカロドントサウルス類と似ている点
まだアロサウルス上科の系統関係が定まっていない時期、例えばアクロカントサウルスやネオヴェナトルがアロサウルス科とされた頃には、シンラプトルがカルカロドントサウルス類と近縁と考えられたことがある。これは主に脳函のためである。シンラプトルの脳函の形態はカルカロドントサウルス類と似ている点が多く、Coria & Currie (2002) によるとCurrie & Carpenter (2000) のデータに脳函の形質を加えて系統解析すると、ギガノトサウルス、カルカロドントサウルスが姉妹群となりシンラプトルを加えた3種がクレードとなった。この3種には5つの共有派生形質があるとしていた。
最も顕著なのは後頭部の角度で、CTスキャンによる脳函の矢状断面をみている。頭蓋天井の前頭骨と、後頭部の面(大後頭孔と基底結節を通る面)のなす角度が、シンラプトル、ギガノトサウルス、カルカロドントサウルスでは鈍角であるが、アクロカントサウルスなど多くの獣脚類ではむしろ鋭角である。これは後頭顆より下で基底後頭骨が後方に突出しているためであるという。またこれら3種では、傍後頭骨突起が強く後側方を向いていることから、後頭顆に対して顎関節の位置がより後方にあったともいっている。
このようなカルカロドントサウルス類との類似は、現在は収斂と考えられている。
(続く)
参考文献
Currie, P. J. & Zhao, X.-J. (1994). A new carnosaur (Dinosauria, Theropoda) from the Jurassic of Xinjiang, People’s Republic of China. Canadian Journal of Earth Sciences, 30 [for 1993], 2037-2081.
Brusatte, S. L. & Sereno, P.C. (2008). Phylogeny of Allosauroidea. Journal of Systematic Palaeontology, 6, 155-182.
Matthew T. Carrano, Roger B. J. Benson & Scott D. Sampson (2012). The phylogeny of Tetanurae (Dinosauria: Theropoda), Journal of Systematic Palaeontology, 10:2, 211-300.
Coria, R. A. & Currie, P. J. (2002). The braincase of Giganotosaurus carolinii (Dinosauria, Theropoda) from the Upper Cretaceous of Argentina. Journal of Vertebrate Paleontology, 22, 802-811.
Wikipedia (English). Metriacanthosauridae.
コメント ( 0 ) | Trackback ( 0 )
中国の
中国のテタヌラ類を調べていて、中国語が読めなくて困った。「額骨」がfrontal 前頭骨だから「前額骨」はprefrontal前前頭骨だろうと類推はできるが、分かりそうで分からないのが辛い。英訳サービスもあるが高すぎる。機械翻訳は避けたい。
例えばレシャンサウルスの論文の中にヘピンゲンシスについてコメントしている部分があるが、読めそうで読めない。。polyglotにもない。
例えばレシャンサウルスの論文の中にヘピンゲンシスについてコメントしている部分があるが、読めそうで読めない。。polyglotにもない。
コメント ( 0 ) | Trackback ( 0 )