獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2016年3月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | ||||
| 6 | 7 | 8 | 9 | 10 | 11 | 12 | ||
| 13 | 14 | 15 | 16 | 17 | 18 | 19 | ||
| 20 | 21 | 22 | 23 | 24 | 25 | 26 | ||
| 27 | 28 | 29 | 30 | 31 | ||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク





| URLをメールで送信する | |
| (for PC & MOBILE) | |
ケラトサウルスのバックル

これはベルトのバックルだそうです。他にアンモナイトもあります。メーカー等はわかりませんが、新宿・紀伊国屋ビルの東京サイエンスにありました。ちょっと高いと思いましたが、アクセサリーとしてはこんなものかな。ケラトサウルス自体が珍しいので買いました。
コメント ( 0 ) | Trackback ( 0 )
ティムルレンギア
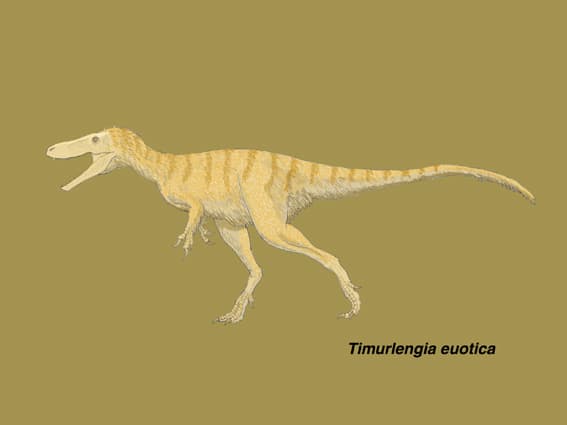

大きい画像
ティラノサウロイドの進化史の中では、ジュラ紀後期から白亜紀前期までの小型で原始的なティラノサウロイドと、白亜紀末に君臨した大型のティラノサウルス科の間にあたる、白亜紀中頃の化石記録がごっそり抜けていた。そのため小型の捕食者から巨大な頂上捕食者に進化する過程で、体の大きさや鋭い感覚などのティラノサウルス科の特徴がどのようにして発達したのかは、よくわかっていなかった。Brusatte et al. (2016) は、そのギャップを埋めるウズベキスタンのBissekty Formation産のティラノサウロイドについて、保存の良い脳函を解析することで、系統的に中間的なティラノサウロイドが存在したことを示している。
結論としてこの恐竜は、まだ体も小さく、頭骨の含気性も発達していなかったが、脳と内耳に白亜紀末の大型ティラノサウルス類にみられる派生的な特徴を持っていた。小型の段階ですでに持っていた脳と鋭い感覚が、頂上捕食者としての進化的成功に役立ったかもしれない、といっている。一部メディアの記事のように、聴覚の発達が体の大型化をもたらした、といった短絡的な表現はしていない。
ティムルレンギアは、白亜紀後期のチューロニアン中期から後期(Bissekty Formation、9000-9200万年前)にウズベキスタンのキジルクム砂漠に生息した中型のティラノサウロイドで、2016年に記載された。
ホロタイプZIN PH 1146/16 は、保存の良い脳函だけである。ただしもう1つの部分的な脳函と、これまでAverianov and Suesによって所属不明のティラノサウロイドと記載されてきた、一連の分離した骨化石も、同一種と考えられている。これらには、左の前頭骨、2個の頸椎、胴椎の神経弓、胴椎、前方の尾椎、中央の尾椎、後方の尾椎、足の末節骨、手の末節骨、右の関節骨と上角骨、左の方形骨、右の歯骨、右の上顎骨が含まれる。これらの骨はティラノサウロイドの特徴をもち、ホロタイプと同じ層準から見つかっており、ほぼ同じ大きさの個体のものと考えられる。
図を見ると、まだまだ断片的な状態である。上顎骨、歯骨とも部分的で、四肢は末節骨しか見つかっていないので、前肢や後肢の長さ、前肢は2本指かどうかなどもわからない。Todd Marshallの美しい全身骨格図がなければ、アマチュアにはつらい論文である。(論文の骨格図は、暫定的にシオングァンロンとティラノサウルスの中間の体形と書いてある。)
ホロタイプの脳函は、シオングァンロンの脳函とほぼ同じ大きさであることから、ティムルレンギアのホロタイプはシオングァンロン(体重170–270 kg)と大体同じ大きさと推定される。全身骨格図では3mちょっとのように見える。脳函の一部で縫合線が閉じていないことから、ホロタイプは完全には成熟していないとも考えられ、成体はもう少し大きい可能性もある。それでも、白亜紀末の大型ティラノサウルス類に比べるとずっと小さい動物と考えられる。
脳のエンドキャストはティラノサウルス科、特にアリオラムスのものと驚くほど似ている。ティムルレンギアには、ティラノサウルス科の脳と内耳の派生的な特徴が多くみられる。たとえば、わずかな中脳屈曲midbrain flexureしか示さないチューブ状の脳エンドキャスト、顕著な中脳頂点midbrain peak、長い蝸牛管などである。(Discussion ではティラノサウルス科の特徴と書いてあるが、Descriptionの中ではティラノサウロイドの特徴と書いてある。)
内耳のエンドキャストをみると、ティラノサウルス科を含めて他のすべての獣脚類と比べても、三半規管と蝸牛管が非常に大きい。これはティムルレンギアの固有形質で機能的意義は不明であるが、敏捷性と関係するのかもしれない。蝸牛管が長いことはティラノサウロイドの特徴で、低周波数の音を聴く能力を高めていた可能性がある。
Brusatte et al. (2016) は、3通りの系統解析を行った。1)ティムルレンギアのデータとして、ホロタイプの脳函だけを用いた場合、2)一連の分離した骨(参照標本)だけを用いた場合、3)ホロタイプの脳函と参照標本の両方を用いた場合、である。その結果、1)2)3)のどの場合でも、細かい違いはあるがティムルレンギアはシオングァンロンに近い段階の中間的なティラノサウロイドと位置づけられた。3)では、ティムルレンギアはシオングァンロンとティラノサウルス科を含むクレードのすぐ外側にきている。
ホロタイプの脳函と参照標本は、別々に解析してもほとんど同じ位置にくること、Bissekty Formationには2種以上のティラノサウロイドが存在したことを示す化石はないことから、同一種である可能性が最も高いという。
ティムルレンギアは、アプティアンからアルビアンの中国のシオングァンロンと多くの類似点をもつ。この2種の脳函は大きさが同じであるだけでなく、よく似たダイアモンド形の上後頭骨の腹側突起をもち、短いbasal tuberaを持っている。さらに、ティムルレンギアの参照標本の1つである長い上顎骨は、シオングァンロンの上顎骨とよく似ているという。シオングァンロン、ティムルレンギア、そしてもしかすると過去にアレクトロサウルスとされたバヤン・シレ産の長い頭骨は、大型で派生的なティラノサウルス類のすぐ外側のグループ(クレードまたは段階)をなすのかもしれないと著者らは考えている。これらの中型で吻の長いタイプの種類の中から、大型のティラノサウルス科が生じてきたのかもしれないという。
(補足)
脳の形については、原始的なティラノサウロイドと白亜紀末のティラノサウルス科のどちらに近いのか、どの部分が進歩的なのかということがポイントと思われるが、原始的なティラノサウロイドと比較した記述はない。ディロングなどの基盤的なティラノサウロイドの脳函は2、3発見されているが、まだCTスキャンもされておらず、詳細な研究はなされていないという。そこはまだこれからということだろう。
参考文献
Stephen L. Brusatte, Alexander Averianov, Hans-Dieter Sues, Amy Muir, and Ian B. Butler (2016) New tyrannosaur from the mid-Cretaceous of Uzbekistan clarifies evolution of giant body sizes and advanced senses in tyrant dinosaurs. PNAS 2016; published ahead of print March 14, 2016, doi:10.1073/pnas.1600140113
コメント ( 0 ) | Trackback ( 0 )
「恐竜博2016」with 宮下さん講演会(2)
恐竜博見学の後は、宮下さん講演会、さらに恐竜倶楽部主催の懇親会と、充実した1日でした。
宮下さん講演会の内容については、多分パンテオンに上がると思いますので、ここでは詳細な紹介はしませんが、現在取り組んでいるいくつかのプロジェクト、ロイヤルティレル及びアルバータ大のフィリップ・カリーに師事した経緯、フィリップ・カリー論、これから研究者を志す若い人たちに向けた座右の銘(ノーベル賞物理学者ワインバーグの4つの心得)からなっていました。
私が最も面白かったのはフィリップ・カリー論の部分ですね。これは、宮下氏がいかにしてアルバータのフィリップ・カリーの元へ押しかけたかという経緯に続いて、カリーの人物像、人柄について語ったものです。宮下氏が10歳の時に読んで恐竜研究を志すきっかけとなった、「最新恐竜ハンドブック」の話。これは筆者も読んだが確かに、他の本と違って「研究する視点」があったと思います。
また2003年の3つの論文、特にCurrie (2003) (北アメリカ産のティラノサウルス類の頭骨の記載・比較解剖学)について、読んでいて心地よいリズムがあるなど絶賛していましたが、宮下氏がいかにフィリップ・カリーに心酔しているかが窺い知れる語り口でした。13年間一緒にいてこれまで一度もフィリップ・カリーに対してがっかりしたことがないそうです。カウディプテリクスなど羽毛恐竜についても主導的な立場にあったが、激化する競争から身を引いて地道な研究に専念したというエピソードについても語っていました。
講演終了後、真鍋先生が、フィリップ・カリー教およびアルバータ教の布教活動とおっしゃったのが受けました。アルバータ教は日本の高校生をアルバータに送り込んでしまう、恐ろしい宗教のようです。
それはいいが、宮下さんの講演を聴いた後で、大変なことに気がついた。
この「肉食の系譜」も初期の段階でフィリップ・カリーの影響を受けているのです。
私theropodが、最初に読んだ恐竜の論文が、宮下さんの絶賛したCurrie (2003)です。ゴルゴサウルスとアルバートサウルスの違いがどうしても知りたかった私は、Currie (2003)にたどり着いた。ポーランドのActa Paleontologica Polonica はオープンアクセスの学術誌で、無料でPDFが入手できた。古生物学の論文なんて、おそらくちんぷんかんぷんだろうと予想していた割には、結構わかりやすく書いてあるので、ある程度は理解できたのです。フィリップ・カリーの論文は、論旨が明確でわかりやすく、余計な部分がない。それでいて他の獣脚類についても豆知識が得られて、教育的でさえある。そのおかげでシンラプトルやシノサウロプテリクス、アクロカントサウルス、マプサウルスなど多くの獣脚類の論文も読む気になった。そうだった。Currie (2003)が素人には到底読めないような難解な文章だったら、「肉食の系譜」もなかった。
私はカリー教の信者ではないと思うが、それは他にもSereno, Xu, Makovicky, Norell など他の研究者の論文に手を広げたからにすぎない。カリー教の魔力は、普通の職業に就いている大人にも作用するのかもしれません。
コメント ( 0 ) | Trackback ( 0 )
「恐竜博2016」with 宮下さん講演会(1)

博物館としては、「恐竜から鳥への進化」のような特定のトピックスや、中国、モンゴル、ゴンドワナといった地域テーマも過去にやり尽くしてきている。一方、世界の恐竜研究は非常に多岐にわたるテーマ、分野で、ものすごい勢いで進捗している。それでも日本の恐竜ファン、恐竜を楽しみにしている子供たちのために、最新の研究成果を紹介する必要がある。それが「7つのキーワード」の背景にあると思われる。とにかく最新研究による新種・新復元・貴重な幼体標本などを一堂に集め、これだけ見られることは大変ありがたいことですね。金曜日にしては混んでいると思いましたが、落ち着いた感じで楽しく観察できる恐竜展でした。
スピノサウルスはまあ、ナショジオにもあったし論文も読んだわけですが、実物化石で足の指の底面が平らなのを確認できました。「スピノサウルス大解剖」のパネルで、複数個体の合成であることがはっきり示されているので、その場にいた女子高生たちも正しく認識していたようで、よかったのではないでしょうか。あとイクチオヴェナトルのデジタル復元も、見ている方はできて当然のようなつもりでも、実際にこれを作成するのはそれなりに大変な作業であるはずで、ありがたいことです。
CG映像のスピノサウルスは、非常に良くできているが骨格図・全身復元骨格と比べると少し後肢が長く作られている。私は見逃しませんよ(笑)。実は私の「スピノサウルスの半水生適応」のイラストも、最初は骨格図を見て描いていたのですが、なんとなく葛藤を感じて後で少し後肢を長くしたのです。同じような心理かもしれないし、将来また覆ったときのための「保険」かもしれません。カプセルフィギュアも明らかに後肢が大きいですね。四足歩行としてもこのぐらいがカッコイイというレベルに調整してある印象です。

スコッティは、スピノサウルスと比較するための恐竜という位置付けですね。しかし今回、左からも右からもゆったりと写真撮影できる配置で、恐竜ファンとしては非常にありがたかった。

ヘテロドントサウルスは何気に珍しいですよね。そうでもないのかな。チレサウルスは意外と気に入った。チリという未知の生態系で全く独特の獣脚類というところがポイント高い。ユニークさの点ではもちろんイーも貴重ですが。

今回一番感動したのは、クリーニング中のサウロルニトレステスです。この化石は、たまりませんね。すごすぎます。従来はヴェロキラプトルと同じくらい細長い顔に描かれてきましたが、もう少しがっしりした顔です。歯もよく保存されていて、やはり前上顎骨は2番目の歯が大きいようですね。maxillary fenestra などの位置は記載されている通りです。下顎から喉のあたりにあるのは舌骨か。SVPのアブストラクトにはあるが、当然ながらまだ研究中で、宮下さんによると論文になるのはいつ頃かわからないそうです。
コメント ( 2 ) | Trackback ( 0 )
ブイトレラプトル


大きい画像
ブイトレラプトルは、白亜紀後期セノマニアンからチューロニアン(Candeleros Formation)にアルゼンチンのリオ・ネグロ州Rio Negro Provinceに生息したドロマエオサウルス類で、2005年に記載された。ゴンドワナに分布したウネンラギア類の中では、最も標本が完全で、最も時代が古いものとされる。というより、Makovicky et al. (2005)によるブイトレラプトルの研究によって、ゴンドワナのドロマエオサウルス類が単系群と認識され、それがウネンラギア亜科 Unenlagiinae と命名された。
ホロタイプには全身のかなりの部分の骨が含まれているが、多くの骨が部分的に失われている。フィールド博物館など海外には復元骨格があるようだが、日本に来たことはないと思われる。
Turner et al. (2012) によると、ブイトレラプトルは次の形質の組み合わせと固有形質により識別される小型のドロマエオサウルス類である。これらのほとんどはMakovicky et al. (2005) と同じである。頭骨が長く、大腿骨の長さよりも25%大きい;歯は小さく、鋸歯がなく、歯冠と歯根の間がくびれていない;方形骨には大きな側方フランジlateral flangeと含気孔がある;後方の頸椎の椎体に腹方外側の稜がある、などである。また他のドロマエオサウルス類と区別される形質として、maxillary fenestra が大きい;前頭骨の縁から後眼窩骨突起までが連続的に移行している(トロオドン類と共有);歯骨に深い歯槽下溝subalveolar groove がある(トロオドン類と共有)、などを追加している。
頭骨はある程度の骨が見つかっているものの、欠損部分が多いので全体の形は推定で復元されている。顔が非常に細長いのは確かで、トロオドン類を通り越して翼竜のような印象もある。一方、歯のついた上顎骨と歯骨は保存されているので、ブイトレラプトルの歯について研究した論文が出ている。
ブイトレラプトルの歯は頭骨のわりに小さく、歯冠の高さが0.6 から 4.6 mmである。歯は後方にカーブし、扁平で、先が非常に尖っている。歯槽に保存された歯も分離した歯も、大きさが異なるだけでみな同じ形をしている。左の歯骨に保存された歯の一つは歯槽の縁が欠けていて、それをみると歯冠と歯根の間が連続的でくびれていない。この点はトロオドン類などと異なる。すべての歯において歯冠の前縁も後縁も少し丸みを帯びていて、鋸歯も稜縁もない。
歯冠の外側(唇側)と内側(舌側)は非常によく似ていて、曲がりがないので唇側と舌側を区別することは困難である。重要な特徴として唇側と舌側の両面に縦のくぼみdepressionがあるため、歯冠の断面が8の字形になっている。さらに、両面に平行に並んだ縦の溝と稜がある。これらの溝と稜は一定のパターンをもつものではなく、数も一定しない。たとえば2本の溝がある場合は稜、溝、稜、溝、稜と並んでいる。
ドロマエオサウルス類の歯の特徴は、より多くの種類が知られるローラシアのドロマエオサウルス類について研究されてきたが、ゴンドワナのウネンラギア類(ブイトレラプトルとアウストロラプトル)は、ローラシアのドロマエオサウルス類とはかなり異なった歯の形態を示すことがわかってきた。Gianechini et al. (2011) は、ウネンラギア類の歯の特徴として1)歯の数が多い、2)歯のサイズが小さい、3)鋸歯も稜縁もない、4)歯冠の表面に縦の溝grooveがある、を挙げている。
1)歯の数が多い:ローラシアのドロマエオサウルス類では下顎の歯骨に11 から16本の歯があるが、ブイトレラプトルでは約25本と推定されている。アウストロラプトルも、少なくとも25本と考えられている。また多くのドロマエオサウルス類は9 から15本の上顎骨歯をもつが、アウストロラプトルは24本である。このような多数の歯はトロオドン類やオルニトミモサウリアのペレカニミムスなどにもみられる。
2)歯のサイズが小さい:歯の相対的な大きさは、上顎骨の最も大きい歯と、上顎骨の丈の高さ(前眼窩窓の前縁の位置で)の比率で表される。この比率はローラシアのドロマエオサウルス類では0.25-0.40 であるが、ブイトレラプトルでは0.18 である。系統解析の結果からするとウネンラギア類の小さい歯は、ローラシアのドロマエオサウルス類にみられる大きい歯が縮小したものと考えられるという。より広くマニラプトル形類をみると、トロオドン類やアヴィアラエ類が0.20より小さく、またテリジノサウルス類エルリコサウルスやアルヴァレスサウルス類シュヴウイアも小さいことから、広く分布する収斂と考えられる。
3)鋸歯も稜縁もない:ローラシアのドロマエオサウルス類では一般に歯の前縁と後縁に鋸歯があるが、ウネンラギア類では前縁にも後縁にも全く鋸歯がない。この鋸歯の分布についてはちょっと複雑である。
ドロマエオサウルスでは前縁にも後縁にも同じくらいの大きさの鋸歯がある。ヴェロキラプトルとデイノニクスでは、前縁の鋸歯が後縁の鋸歯よりずっと小さい。サウロルニトレステスも同様だが、この種では前縁の鋸歯があるものとないものがみられる。ミクロラプトルの前上顎骨歯と前方の歯骨歯には鋸歯がない。シノルニトサウルスでは、前上顎骨歯、最初の上顎骨歯、前方の歯骨歯には全く鋸歯がなく、その後方の一部の歯では後縁だけに鋸歯がある。ドロマエオサウルス類では、一部の歯で前縁の鋸歯を失っているのが基本で、その後二次的に前縁の鋸歯を回復してきたと考えられているらしい。(それでドロマエオサウルス亜科が派生的となるようである。)その中にあって、完全に鋸歯がないことはウネンラギア類の共有派生形質と考えられるという。
4)歯冠の表面に縦の溝grooveがある:ウネンラギア類の特徴の一つは歯冠に縦の溝があることである。この溝は、系統的には離れているがスピノサウルス類やケラトサウルスにみられるものと若干似ている。El Anfiteatroという場所で発見された分離した歯、Endemas-Pv 15はウネンラギア類と比較されるマニラプトル形類とされている。この歯は鋸歯がなく、強く後方に曲がっており、断面が8の字形であり、またエナメルのひだfoldがある点で、ブイトレラプトルと似ている。ただし大きさがブイトレラプトルよりずっと大きく、19 mm もあることから、別のウネンラギア類と考えられる。他にもドロマエオサウルス類の可能性がある歯で、溝や稜のあるものがモロッコ、スペイン、フランスなどから報告されているが、これらは鋸歯があるという。
Gianechini et al. (2011) は、縦の溝のある歯はスピノサウルス類、ノアサウルス類、モササウルス類、翼竜など、魚食性の四足動物にみられることは興味深いといっている。ブイトレラプトルもアウストロラプトルも、豊富な魚類化石を含む河川堆積物の近くで発見されていることから、魚食性の可能性があると述べている。このスコミムスか偽歯鳥類のような長い吻を、水面下に突っ込んで魚類などを捕食したのだろうか。
これほど特殊化した動物が、ドロマエオサウルス類の中で基盤的な位置にくる理由は何だろう。Makovicky et al. (2005) によると肩帯、腰帯、後肢などに系統上重要な形質があるようだ。烏口骨は直角に曲がっているが、これはシノルニトサウルスと似ている。上腕骨は肩甲骨よりも30%長いが、これは鳥類と遼寧省のドロマエオサウルス類(ミクロラプトル類)にしかみられない。腸骨には顕著なsupra-acetabular crestがあり、これはウネンラギアやラホナヴィスと同様である。座骨の閉鎖突起obturator processの形は、ラホナヴィス、ミクロラプトル、シノルニトサウルスと似ている。中足骨はアルクトメタターサルで、尾椎の前関節突起はあまり長くない。これらはドロマエオサウルス類としては原始的な形質ということかもしれない。
参考文献
Gianechini, F.A., Makovicky, P.J., and Apesteguia, S. (2011). The teeth of the unenlagiine theropod Buitreraptor from the Cretaceous of Patagonia, Argentina, and the unusual dentition of the Gondwanan dromaeosaurids. Acta Palaeontologica Polonica 56 (2): 279-290.
Makovicky, P.J., Apesteguia, S., and Agnolin, F.L. (2005). The earliest dromaeosaurid theropod from South America. Nature 437: 1007-1011.
Alan H. Turner, Peter J. Makovicky, and Mark A. Norell (2012) A Review of Dromaeosaurid Systematics and Paravian Phylogeny. Bulletin of the American Museum of Natural History, Number 371:1-206.
コメント ( 0 ) | Trackback ( 0 )