獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2024年12月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | 31 | ||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
ケラトサウルス・ナシコルニス

ケラトサウルスのホロタイプ標本USNM 4735は、スミソニアンの国立自然史博物館に長年、展示されていたが、改装工事のため取り外され、再クリーニングが進められた。その機会にCarrano & Choiniere (2016)は、ケラトサウルスの前肢の骨格について研究し、アベリサウルス類などケラトサウリアの前肢の退化との関連についても考察した。ホロタイプ標本では左の前腕と手がよく保存されている。ただし指骨は完全ではないので、正確な指骨の数はわからない。
ケラトサウルスの前肢は関節状態で堆積したと考えられるが、手根骨は保存されていない。前腕(橈骨、尺骨)と手(中手骨、指骨)の間に隙間があるので、軟骨性の手根骨が1,2個あったと考えられている。橈骨・尺骨はディロフォサウルスやエオアベリサウルスと似ている。また中手骨もディロフォサウルス、ベルベロサウルス、エオアベリサウルスと似ており、特殊化したリムサウルスとは似ていない。中手骨はI~IVの4個とも保存されており、中央のII, III が大きくI, IVは小さい。指骨はII, III, IV に1個ずつ保存されている。第I指の指骨は保存されていないが、中手骨の関節面から指骨があったとわかる。第II, III指にはII-1, III-1が保存されており、関節面があることから短いが機能的な指があったと考えられる。第IV指には1個だけ小塊状の指骨があるが、半球状の関節面があることから、あと1個指骨があったことが示唆される。痕跡的ながら2個あったということである。
ケラトサウリアの中の系統関係は、研究者の間で一致していない。ケラトサウリアにはケラトサウルス科、エラフロサウルス科、ノアサウルス科、アベリサウルス科というべき系統があり、伝統的にはケラトサウルス科が基盤的で、ノアサウルス科とアベリサウルス科が互いに近縁で合わせてアベリサウルス上科Abelisauroideaをなす、と考えられてきた。
Wang et al. (2017) はリムサウルスの個体発生の論文で、ケラトサウリアの大規模な系統解析を行った結果、まずエラフロサウルス類を含むノアサウルス類と大型のケラトサウルス科やアベリサウルス科を含むグループに分かれるという系統関係を得た。つまりケラトサウルス科とアベリサウルス科はより近縁となった。
Delcourt (2018)はケラトサウリアの短い総説の中で、Wang et al. (2017)の系統解析を採用し、ケラトサウルス科とアベリサウルス科を合わせたクレードをエトリガンサウリアEtrigansauria と命名した。ここではケラトサウルス科にエオアベリサウルス、ケラトサウルス、ゲニオデクテスが含まれている。
しかしその後もアルゼンチンのアベリサウルス類の研究者は、伝統的な系統関係を変えておらず、Wang et al. (2017) やDelcourt (2018)の考えが定着しているわけでもなさそうである。
参考文献
Matthew T. Carrano & Jonah Choiniere (2016) New information on the forearm and manus of Ceratosaurus nasicornis Marsh, 1884 (Dinosauria, Theropoda), with implications for theropod forelimb evolution, Journal of Vertebrate Paleontology, 36:2, e1054497, DOI: 10.1080/02724634.2015.1054497
Rafael Delcourt (2018) Ceratosaur palaeobiology: new insights on evolution and ecology of the southern rulers. Scientific Reports (2018) 8: 9730 | DOI:10.1038/s41598-018-28154-x
Wang, S. et al. (2017) Extreme Ontogenetic Changes in a Ceratosaurian Theropod. Curr. Biol. 27, 144–148.
コメント ( 0 ) | Trackback ( 0 )
サルトリオヴェナトル(3)
感想であるが、恐竜の指が外側から順に失われてきたことは従来からいわれていたことであり、またサルトリオヴェナトルの手も不完全なので、それが決定打といえるかどうかはともかく、ケラトサウルスの指が完全には見つかっていないのを補った形になっている。他の種類とともにケラトサウリアの祖先形を推定するのに役立っているわけである。こうしてサルトリオヴェナトル、ケラトサウルス、エオアベリサウルス、リムサウルスと並べてみると、リムサウルスの手は同じジュラ紀のエオアベリサウルスと比べても退化的で、特殊化したものであることがわかる。これをもとにアヴェロストラの祖先形やテタヌラ類のことを論じるのは、やはり無理があると思える。
そもそもテタヌラ類が3本指といっても、基盤的なテタヌラ類では第IV中手骨が残っているわけで、普通に考えると(コエロフィソイドで既に第IV指が退化的であることなどを考えると)I-II-III-IV で、これをII-III-IV-V とみるのは難しいのではないか。
ここではメトリアカントサウルス科は解体されて、シュワンハノサウルスやスゼチュアノサウルス(ヤンチュアノサウルス)・ジゴンゲンシスは基盤的な位置にきている。ティラノサウルス類の中でというのは、グァンロンとメガラプトルで第IV中手骨が残っているためである。アロサウロイドの中では、シンラプトルで第IV中手骨が残っているので、アロサウリア(アロサウルス以上)で退化した、ということである。コエルロサウルス類の中ではティラノサウロイドが分岐した時点では残っていたはずなので、マニラプトル形類の祖先で退化したことになる。
ただしサルトリオヴェナトルが断片的であることを考えると、もし頭骨や脊椎が見つかってサルトリオヴェナトルが実はテタヌラ類やディロフォサウルス類になったりすると、このままのロジックでは使えなくなるということだろう。
恐竜が好きな人なら、コレクタ社のフィギュアを眺めるだけでも楽しめる。なかなかよくできている。しかし前肢を見て、この第III指が完全に保存されていることが、重要な意義があるのだなと思いをはせれば、さらに楽しめる。
コメント ( 0 ) | Trackback ( 0 )
サルトリオヴェナトル(2)
サルトリオヴェナトルの手は、図のように第II 中手骨と指骨II-1、第III指の指骨すべてがほぼ完全に保存されている。この部分的に保存された手の情報と、系統解析の結果、基盤的なケラトサウリアになったことから、テタヌラ類の3本指の進化についての議論に貢献している。
詳しい方はご存知と思うが、この議論とは鳥の前肢の3本指の相同性についての論争である。現生鳥類の3本指は、I, II, III であるのか、 II, III, IVであるのかという問題である。元々、古生物学と比較解剖学の知見からは、恐竜の手の指は5本指の祖先から第V指、第IV指の順に退化して、テタヌラ類の3本指になったことは明らかに思われた。鳥類に近いデイノニクスやアルカエオプテリクスに至るまで、問題なくI, II, IIIと考えられた。ところがニワトリ胚などの発生学的研究からは、II, III, IVと考えられた。四肢動物では指の原基となる中胚葉細胞の凝集塊が、枝分かれしたパターンをなすが、その主軸と呼ばれるメインの軸が、ワニやカメなど含めてどんな四肢動物でも第IV指を通っている。そのパターンからはニワトリ胚の3本指はII, III, IVと考えられたわけである。この矛盾はかつて、鳥類の祖先は恐竜ではないという説の重要な根拠とされていた。
Xu et al. (2009) はノアサウルス類リムサウルスの手の指のパターン(指が退化的な4本指)から、3本指の獣脚類でも発生学が示唆するようにII, III, IVであると提唱した。リムサウルスの手は4本指の両端の指が退化的であるが、Iの方が先行して退化しているようにみえる。彼らはリムサウルスを基盤的なケラトサウリアと考え、テタヌラ類の祖先でもII, III, IVが残り、よってテタヌラ類の3本指もII, III, IVであると考えた。(実際にXuらのグループの研究では現在もそのように表記している。)

Copyright 2018 Dal Sasso et al.
Dal Sassoらは系統解析の結果から、テタヌラ類の3本指に至る各系統の祖先形の指骨式を推定した(図)。恐竜全体の祖先では、ヘテロドントサウルスのように2-3-4-3-2である。竜盤類、獣脚類、新獣脚類の祖先では、コエロフィシスのように2-3-4-1-0となる。アヴェロストラ(ケラトサウリア+テタヌラ)の祖先形は2-3-4-1-Xとなった。テタヌラ類の祖先では、2-3-4-0-Xである。ここで0とは、指骨が失われて中手骨は残っている状態をさす。Xとは、中手骨さえも失われた状態をさす。鳥類を含む進化したテタヌラ類では2-3-4-X-Xである。
ケラトサウリアでは、エオアベリサウルスの第I指、リムサウルスの第II指、サルトリオヴェナトルの第III指が完全に保存されていること、またリムサウルスやマジュンガサウルスでも第IV指に1個の指骨があることから、ケラトサウリアの祖先形は2-3-4-1-Xと推定される。またサルトリオヴェナトルの第II中手骨の形態は、基盤的なケラトサウリアの第II中手骨とも、基盤的なテタヌラ類(アクロカントサウルス、スゼチュアノサウルス・ジゴンゲンシス、シュワンハノサウルス)の最も長い中手骨とも似ており、これらの間のギャップを埋めている。このことからテタヌラ類の最も長い中手骨は第II中手骨であると確認される。またサルトリオヴェナトルの第III指は、コエロフィソイドなどの第III指とも、テタヌラ類の最も外側の指ともよく似ており、これらの相同性を強く支持している。
リムサウルスの状態(0-3-3-1-X)は、ケラトサウリアの祖先形と比べて大きく派生したもので、3本指のテタヌラ類の祖先形とは考えられないとしている。
基盤的なテタヌラ類には痕跡的な第IV中手骨が残っていることから、テタヌラ類の祖先形は2-3-4-0-Xと考えられる。ケラトサウリアでは手が退化したアベリサウルス類でも第IV中手骨が残っていることから、発生過程で主軸の位置は第4指のまま変わらなかったと思われる。それに対してテタヌラ類では、第IV中手骨の退化が独立していくつかの系統で起きている(アロサウルス類、ティラノサウルス類、マニラプトル形類)ことから、テタヌラ類の祖先のどこかで、第4指から第3指の位置へ主軸の移動が起きたことを示唆している。結局、鳥類の3本指の進化は、祖先の恐竜において外側の指から順に失われ、第IV指が完全に失われたテタヌラ類の祖先の段階で、主軸の移動が起きたと考えるのが最も説明しやすいと述べている。
コメント ( 0 ) | Trackback ( 0 )
サルトリオヴェナトル(1)

コレクタ社は時々、変なものを作るが、その中には恐竜ファンにとって貴重なものもある。群馬の翼竜展にあったカビラマスというフィギュアは、初期の翼竜ラエティコダクティルスの全身復元であり、売れ行きはどうか知らないが、貴重なものである。コレクタ社の商品には、ヨーロッパでの重要な発見が反映された、ヨーロッパの恐竜ファンにとって思い入れのある種類が含まれているということだ。2020年の新作の「ソルトリオベネーター」は、おとなしめのケラトサウルスみたいなものであるが、誰がこんなマイナーなものを買うのかと思ったら、結構売れているのか在庫なしとなっている。これがどういう恐竜か、皆がよくわかって買っているとも思えない。
しかし長年の恐竜ファンなら、ピンとくる。昔、「ディノプレス」にイタリアで発見された「サルトリオサウルス」が載っていた。初期のテタヌラ類ということで、シンラプトルそっくりのイラストがあったのである。ああ、あれがケラトサウリアになったのだなとわかる。
1996年、アマチュア化石コレクターでミラノ自然史博物館とも協力しているAngelo Zanella氏が、イタリア北部のサルトリオ近くのアルプス山麓の採掘場で、岩のブロックにいくつかの骨を発見し、ミラノ自然史博物館に知らせた。化石を含む地層は鉱業用の爆薬で何百もの破片に砕かれていたため、その後の採集は困難だったという。サルトリオは15世紀以来、良質な大理石の産地として有名だったらしい。1999年、1800時間に及ぶ膨大な化学的プレパレーション作業の末に、132個の化石片が分離された。下顎の断片、肋骨の断片、肩帯や四肢の骨が同定され、大型獣脚類のものとわかった。この標本はサルトリオサウロ“saltriosauro”の名で有名となり、海外ではラテン語化されたサルトリオサウルスと呼ばれたが、正式に記載されていないので有効な学名ではなかった。これがDal Sasso et al. (2018)によってサルトリオヴェナトル・ザネライSaltriovenator zanellaiとして記載された。サルトリオヴェナトルは、ジュラ紀前期では最大の獣脚類と考えられ、イタリアではスキピオニクスに次いで2番目の獣脚類となった。
サルトリオヴェナトルは、ジュラ紀前期シネムリアン(Saltrio Formation)にイタリア北部のロンバルディア州に生息した基盤的なケラトサウリアで、2018年に記載された。(最も基盤的なケラトサウリアであってケラトサウルス科ではないが、このサイトでは便宜上ケラトサウルス科のカテゴリーに収録している。)サルトリオヴェナトルの骨格には、4本指の獣脚類(コエロフィソイドやケラトサウリア)と原始的なテタヌラ類の特徴がモザイク的に混じっているという。
サルトリオヴェナトルのホロタイプ標本は断片的な部分骨格で、下顎の断片(部分的な右の板状骨と右の前関節骨)、頸椎と胴椎の肋骨、叉骨、左の肩甲骨、右の肩甲骨の関節窩、右の烏口骨、右の胸骨の断片、右上腕骨、部分的な左上腕骨、部分的な右手(手根骨、中手骨、指骨)、部分的な右足(足根骨と中足骨)からなる。その他に、骨の近くに1本の分離した歯が見つかっていて、獣脚類の上顎骨または歯骨の歯であるが、歯根がなく顎との関係もわからないため、慎重にホロタイプからは除外して参照標本となっている。

Copyright 2018 Dal Sasso et al.
発見された骨の部位を表す図が非常によくできていて、下顎の内側が見えるようになっているし、左右の肩帯、前肢、後肢も同時に見えるように描かれている。さらに、このアングルだと見つかっている部分が多めに見える。復元された全身像はやたらかっこいいが、これはケラトサウルスの良さだろう。見つかった部分は断片的なので、正確な全身復元は不可能なのがわかる。それでも著者らは、全身骨格の復元と全長の推定を試みている。
系統解析の結果、サルトリオヴェナトルはモロッコのベルベロサウルスと姉妹群をなし、最も基盤的なケラトサウリアと位置づけられた。サルトリオヴェナトルが他のケラトサウリアと共有する派生形質は、太い第II中手骨(長さが遠位の幅の5/2を超えない)、第II中手骨のcollateral ligament fossaにはっきりした棚状の縁がある、などの非常に細かい手の形質である。
サルトリオヴェナトルを強制的にテタヌラ類に入れようとした場合は、分岐樹が5ステップ長くなってしまう。その場合にはサルトリオヴェナトルは、最も基盤的なテタヌラ類となったり、スゼチュアノサウルス・ジゴンゲンシスと姉妹群になったり、コエルロサウルス類となったりしたという。
サルトリオヴェナトルの肩甲骨と上腕骨は、全長8mのアロサウルスの亜成体(MOR 693)の骨格と比べて、それぞれ同じか10%大きかった。後肢の足根骨と中足骨は、両者で同じくらいの大きさだった。サルトリオヴェナトルの前肢の骨は、MOR 693よりも太くがっしりしていた。これらのことから、サルトリオヴェナトルの全長は7~8 mと推定された。
これまではクリオロフォサウルスが前期ジュラ紀最大の獣脚類とされていた。クリオロフォサウルスはケラトサウルスの大型の標本UMNH 5278と同じくらいの大きさと考えられているが、サルトリオヴェナトルはこれよりも大きいという。
Dececchi & Larsson (2011)の定量形態学的データによると肩甲骨または上腕骨の長さと大腿骨の長さには強い相関があり、その計算式に基づいて計算すると、サルトリオヴェナトルの大腿骨は822 ~887 mmで、これはクリオロフォサウルスよりも5–13% 大きいという。この大腿骨長から体重を推定すると、1,269–1,622 kg となった。
サルトリオヴェナトルは基盤的なケラトサウリアとなったので、その全身骨格の復元には最も完全に知られるケラトサウルスの骨格をベースとして、サルトリオヴェナトルの保存された骨を、サイズを合わせて当てはめていった。見つかっていない部分については系統上、ケラトサウリアの共通祖先において推定される状態を描いたという。どうみてもケラトサウルスに見えるが、鼻骨の正中のとさかや涙骨の突起は、最も基盤的なアヴェロストラ、つまりケラトサウリア(ケラトサウルス)+テタヌラ(モノロフォサウルス、スピノサウルス類、プロケラトサウルス類)で様々に発達している。そのためサルトリオヴェナトルの復元にも描かれている、とちゃんと述べている。
体の輪郭・肉付けについてはグレゴリー・ポールなどの筋肉の配置に基づき、得られた全身像は頭骨が80 cm、全長730 cm、腰の高さ220 cmとなった。大腿骨長が800–870 mmとなり、体重は1,160–1,524 kgとなった。
いずれにしてもサルトリオヴェナトルは、体重1,000 kgに近いサイズであり、ジュラ紀前期としてはクリオロフォサウルスをしのぐ最大の肉食恐竜となった。これはジュラ紀後期のシンラプトルやアロサウルスに匹敵するものであるという。
参考文献
Dal Sasso C, Maganuco S, Cau A. (2018). The oldest ceratosaurian (Dinosauria: Theropoda), from the Lower Jurassic of Italy, sheds light on the evolution of the three-fingered hand of birds. PeerJ 6:e5976 DOI 10.7717/peerj.5976
コメント ( 0 ) | Trackback ( 0 )
ゲニオデクテス1

ゲニオデクテスは、今から百年以上も前にアルゼンチンのパタゴニアで発見された謎の肉食恐竜で、1901年にWoodwardによって記載された。吻の一部、つまり大きな歯の付いた上顎と下顎しか見つかっていないので、その全身像は現在でも不明のままである。断片的な標本であるため、メガロサウルス科とされたりティラノサウルス科とされたり、またアベリサウルス科に似ているともいわれてきたが、2004年にRauhutにより再記載された。それによると、ケラトサウルスと最も近縁であるという。また化石の正確な産出地と年代が不明であったが、おそらくチューブート州のカニャドン・グランデのセロ・バルシノ層で、白亜紀前期アプト期からオーブ期と考えられるという。
おそらくゲニオデクテスとケラトサウルスだけに共通する特徴として、前上顎骨歯が互いに重なった梯形に並んでいることと、最も長い上顎骨歯の歯冠が下顎(歯骨)の最も細い部分の丈よりも長いことがあげられる。ゲニオデクテスとケラトサウルスが異なる点は、ケラトサウルスでは前上顎骨歯が3本であるのに対して、ゲニオデクテスでは4本あることである。
ゲニオデクテスの前上顎骨は比較的がっしりしていて、吻の先端は腹側からみて幅広いU字型をしている。鼻孔の下の体部は、長さと高さが同じくらいであり、4本の歯を備えている。上顎骨は歯の生えた部分だけが保存されているが、残っている部分には前眼窩窩の腹側縁は含まれていないことから、それは比較的高い位置にあったと考えられる。上顎骨歯が長いので歯槽も深いということだろう。また上顎骨の側面は、少数の孔以外は平滑だったらしいという。下顎の歯骨はややがっしりしていて、歯骨の前端が背側に少し膨らんでいるので、3番目より後ろの歯列が凹型にカーブしている。歯骨の側面では、5番目の歯槽から後方に、歯槽側の縁から高さの1/3の距離に溝がある。前上顎骨、上顎骨、歯骨のいずれにおいても、歯間板は癒合している。
前上顎骨以外は正確な歯の数はわからないが、他の獣脚類との比較から推察すると、上顎骨歯は15本以下、歯骨歯は14本以下と考えられる。他の多くの獣脚類では長い歯と成長中の小さい歯が交互に並んだパターンがみられるが、ゲニオデクテスではほとんどの歯が同調して成長しているようにみえ、成長中の小さい歯は少ない。前上顎骨歯は上顎骨歯よりも顕著に短い。上顎骨歯はかなり長いが、これは歯槽から抜けかけているというようなアーティファクト(人工産物)ではなく、実際に長いようである。後縁の鋸歯がほとんど歯槽の縁まで存在することや、歯冠と歯根の境界がどの歯にもみられないこと、さらに上顎骨の浸食された所をみると、歯根が歯槽にフィットしていることから、歯の位置が大きくずれてはいないと考えられる。最も長い上顎骨歯は復元すると95 mmに達し、下顎の最も細い部分の高さよりも長い。上顎骨歯は非常に強く側扁している。また歯冠の前縁と後縁に沿って,特徴的な平らな部分がある。
ゲニオデクテスでは、前上顎骨歯が上顎骨歯よりもかなり小さいが、これはティラノサウルス科の特徴の一つである。しかしゲニオデクテスの前上顎骨歯は、ティラノサウルス類と異なり断面がD字型ではない。上顎骨歯の形状もティラノサウルス類とは全く似ていない。さらに、前上顎骨歯と上顎骨歯の大きさの顕著な違いは、ケラトサウルスにもみられる。
上顎骨歯の辺縁に沿って平らな領域が存在するという特徴は、ケラトサウルスとアベリサウルス類にみられる。この領域は他の獣脚類では通常凸型であることから、この特徴はネオケラトサウリアの共有派生形質と考えられる。完全に癒合した歯間板は、ネオケラトサウリアを含めていくつかの系統にみられるが、総合的に考えるとこれもネオケラトサウリアの形質とみるべきだろうという。
ゲニオデクテスの上顎骨歯の特徴の一つは歯冠が非常に扁平なことであり、このように扁平な歯は他にはケラトサウルスとカルカロドントサウルス類にしかみられない。しかしカルカロドントサウルス類の歯は、鋸歯の基部にある下向きの溝やエナメルのしわがある点で,ゲニオデクテスとは異なる。
ケラトサウルスと共通するもう一つの特徴が,上顎骨歯が非常に長いことである。ケラトサウルス・デンティスルカトゥスでは最も長い上顎骨歯の歯冠が93 mmで、歯骨の最も細い部分の丈に匹敵する。ケラトサウルス・ナシコルニスでも70 mmで、歯骨の最も細い部分の丈より長い。標本が断片的なため得られる情報は限られているが、以上のことからゲニオデクテスはケラトサウルス科(ネオケラトサウリアのうち、アベリサウルス類よりもケラトサウルスに近縁)と結論している。
参考文献
Rauhut, O. W. M. 2004. Provenance and anatomy of Genyodectes serus, a large-toothed ceratosaur (Dinosauria: Theropoda) from Patagonia. Journal of Vertebrate Paleontology 24: 894-902.
コメント ( 0 ) | Trackback ( 0 )
ケラトサウルス1
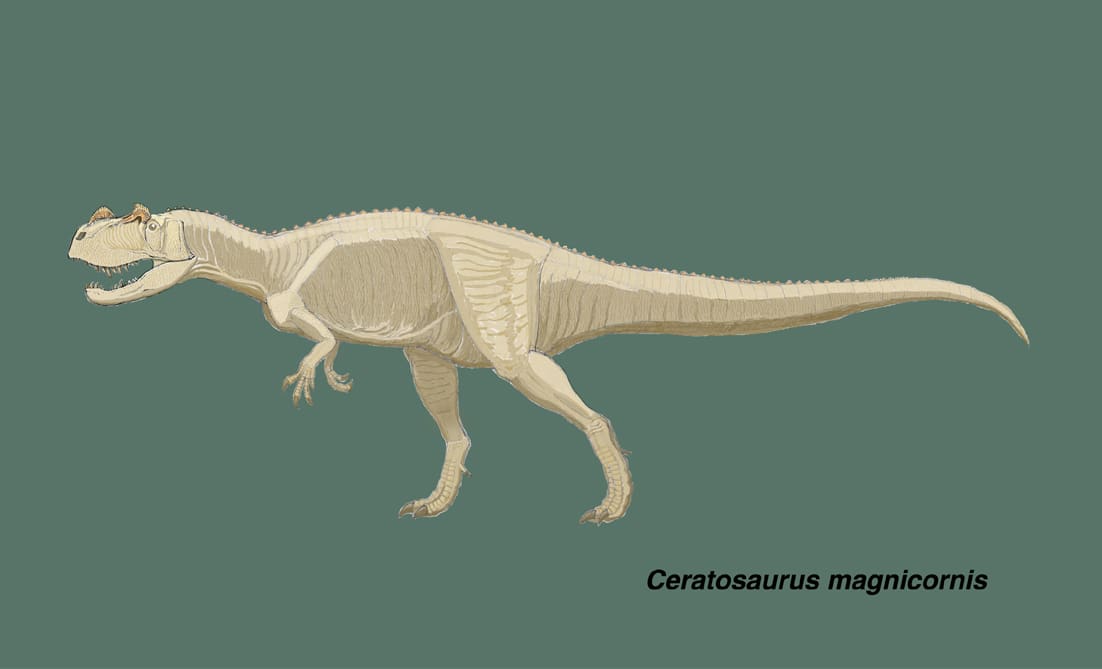
ケラトサウルスはジュラ紀後期に北アメリカ、アフリカなどの地域に生息した獣脚類で、1884年にマーシュにより記載され,1920年にギルモアによって再記載された。長年にわたり単一の種ケラトサウルス・ナシコルニスのみが認められていたが、2000年にマドセンとウェレスによって2つの新種が記載された。ユタ州クリーブランド・ロイド発掘地から発見されたケラトサウルス・デンティスルカトゥスと、コロラド州西部から発見されたケラトサウルス・マグニコルニスである。
ケラトサウルス・マグニコルニスは、部分的に関節したほとんど完全な頭骨を含む保存のよい骨格がみつかっている。頭骨はナシコルニスのそれと比べて、全体にがっしりしていて、より長く丈が低い。上顎骨は長く,その下縁はより丸くカーブしている。上顎骨には13本の歯があり、ナシコルニスよりも長く太い。上顎骨の鼻骨突起には洋なし形の穴maxillary sinusがある。涙骨の角は半円状で、周縁部に放射状の溝が走っている。鼻骨の角はナシコルニスよりも前後に長く,やはり放射状の溝がある。前上顎骨には3本の歯があり、梯形に並んでいる。ケラトサウルスの特徴として前上顎骨歯と最初の3本の歯骨歯には、歯の内側(舌側)に縦の溝がある。下顎の歯骨は上向きにカーブしていて下顎の先端が丸くなっている。(このマグニコルニスの歯骨については、全般的特徴の項には具体的なことが書かれているのに、個々の骨の詳細な記載の項にはなく、図版もなく、最後の標本リストにもない。どういう事情なのだろうか。)
ケラトサウルス・デンティスルカトゥスは3種の中では最も大型で、肋骨や血道弓なども含む全身の骨が発見されている。ただし頭骨については前上顎骨、上顎骨、方形骨、方形頬骨、歯骨、角骨、板状骨など、上顎と下顎周辺の骨しかなく、涙骨や鼻骨が見つかっていないので角の正確な形状はわからないはずである。ケラトサウルス・ナシコルニスでは脊椎に沿って、皮骨の小塊dermal ossicleが並んでいたとされ、2つの新種でもこの皮骨の小塊がみつかっている。
1925年にヤネンシュは、タンザニアのテンダグルから発見された断片的な標本をもとにケラトサウルス・レヒリンギを報告したが、マドセンらは種レベルの同定はできないとしてケラトサウルスの一種(Ceratosaurus sp.)としている。この方形骨の大きさはマグニコルニスの倍近くもあり、また特徴的な縦の溝のある歯も見つかっていることから、アフリカにはケラトサウルスの大型種が生息していたことは確かであるという。
ケラトサウルスは眼の上、鼻の上に角をもち、短剣状の歯は長い、背中に皮骨が並んでいるなど、けっこう派手な肉食恐竜といえる。ナシコルニスでは鼻骨の角がわりと尖っているようであるが、マグニコルニスでは丸みを帯びている。骨芯の上に皮膚の角質が覆うので、生体復元では骨芯よりもかなり大きく見えるわけであるが,骨芯が丸いので角質も丸みを帯びた感じにしておいた。ケラトサウルスは上顎の歯がかなり長い。最も長い何本かは抜けかけの状態かもしれないが、実際に歯が上顎についた状態で見つかっているのであるから、そのように復元しなければならないだろう。これをどの程度上唇で覆うかで印象が変わってくる。アロサウルスでいわれているように、捕食に際してサーベルタイガー的な使い方をしたのであろうか。
参考文献
Madsen, J. H., and S. P. Welles. 2000. Ceratosaurus (Dinosauria, Theropoda). A revised osteology. Miscellaneous Publication, Utah Geological Survey 00-2:1-80.
コメント ( 0 ) | Trackback ( 0 )