獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2024年12月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | 31 | ||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
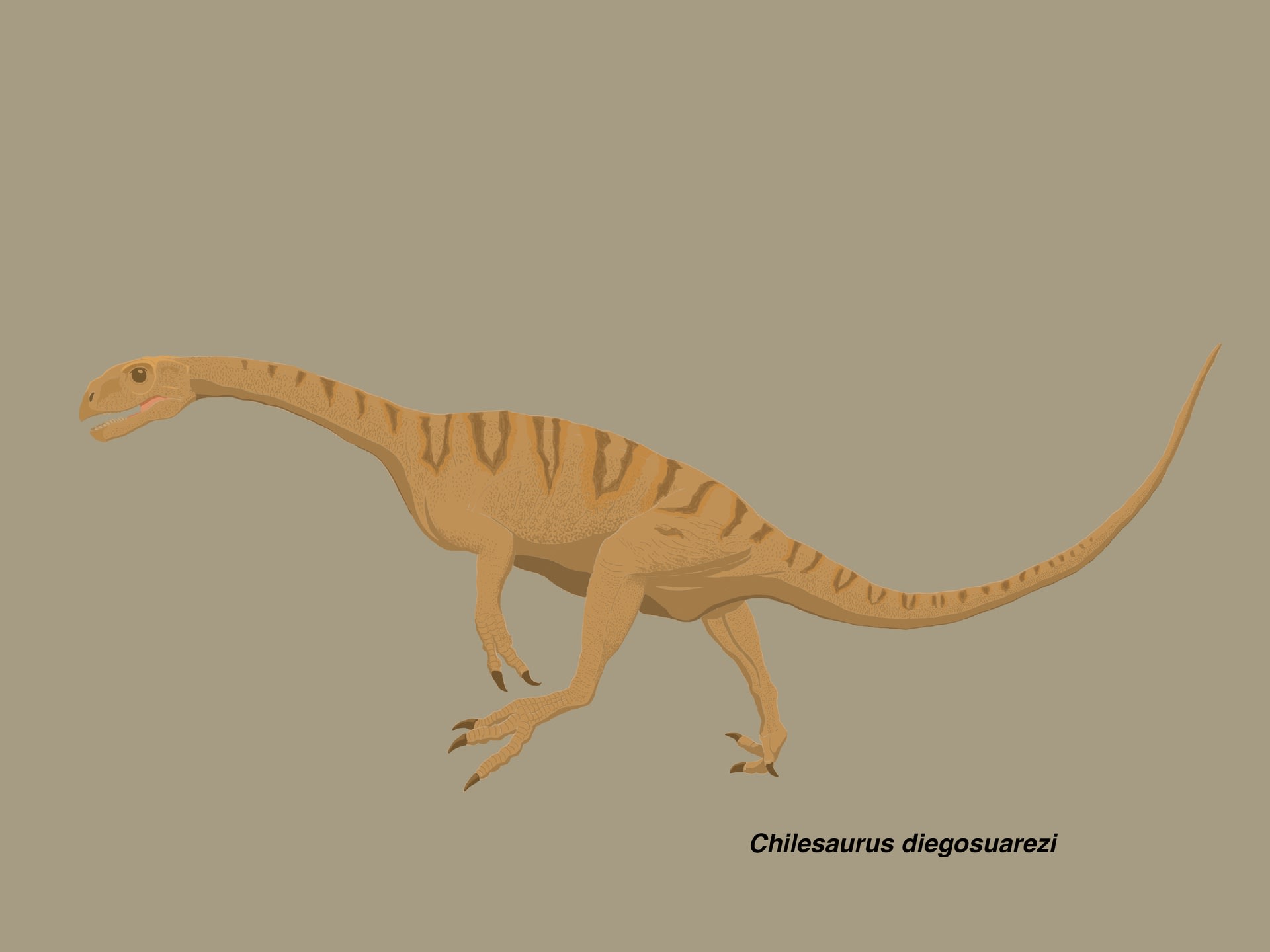
チレサウルス
また非常にユニークな、面白い恐竜が現れた。獣脚類全体の進化史を考える上で、重要な種類にちがいない。
チレサウルスは、ジュラ紀後期チトン期(Toqui Formation)に南米のチリ南部に生息した基盤的テタヌラ類で、2015年にNovas et al.により記載された。植物食に適応した歯や腰帯、後肢などには、原竜脚類、テリジノサウルス類、はては原始的鳥盤類とも似た特徴がモザイク的に入り混じっているが、系統解析の結果から獣脚類とされた。ちなみに、一部ネットメディアの見出しのような「ティラノサウルスの近縁種」ではない。2本指であることはティラノサウルスとの類縁とは直接関係がない。前肢の指の数が減る傾向は獣脚類らしいといえばそうかもしれないが、もちろん主な根拠ではない。
チレサウルスのホロタイプ(SNGM-1935)は完全に近い、関節した全身骨格で全長約1.6 mと推定されるが、これはまだ成熟していない亜成体で、最も大きい標本の50%の大きさである。パラタイプは成長段階の異なる4個体の部分骨格で、全長1.2 から 3.2 mであるという。
チレサウルスに固有の特徴は、歯が葉状 leaf-shaped で萌出中の歯の歯冠の頂端にのみ鋸歯がある、烏口骨が四角形に近く厚い縁をもつ、手の第2指の末節骨の手前の指骨が短い、第3指が退化している、恥骨が完全に後方を向いている、恥骨の軸が棒状 rod-like で遠位端がふくらんでいない、などである。
頭骨は、分離した前上顎骨、上顎骨、前頭骨、後眼窩骨、歯骨などが見つかっており、頭は比較的小さかったと考えられる。前上顎骨は短く丈が高く、前端に粗面があり角質のくちばしがあったらしい。歯骨は前端の方が丈が高いので、テリジノサウルス類のように腹側縁が下方にカーブしている。歯骨の歯は丈が高く、葉状で、前方に傾いておりprocumbent、歯冠の頂端にのみ小さな鋸歯がある。
頸椎は長く、細長い首をなす。頸椎と前方の胴椎には、一対の仕切りのある側腔 pleurocoelがある。
四肢の骨は原竜脚類のようにがっしりしている。上腕骨は短く幅が広い。中手骨は第1から第3まであるが、よく発達しているのは第1指と第2指のみである。第1指の末節骨は他のテタヌラ類ほどカーブしていない。第2指の末節骨の手前の指骨が強く短縮している。厳密には2本指ではなく、第3指には中手骨だけではなく、1個の小さい指骨がある。(これは論文の写真には示されておらず、supplementary informationにもない。)
腸骨には竜脚形類、テリジノサウルス類、コエルロサウルス類、いくつかの鳥盤類と共通の特徴がある。恥骨は鳥盤類、テリジノサウルス類、ドロマエオサウルス類と同様に、完全に後方を向いている。恥骨の先端がふくらんでいないことは、多くの獣脚類とは対照的である。
足根部(足首)は基盤的竜盤類に似ている。距骨の上方突起は低く太く、板状で高く伸びているテタヌラ類とは異なる。ヘレラサウルスや基盤的竜脚形類のような初期の竜盤類と同様に、チレサウルスの踵骨は遠位側からみて三角形に近い。獣脚類の踵骨は長方形である。
チレサウルスの第1中足骨は第2中足骨の50%以上あり、これは他の多くの獣脚類と異なる。足の第1指は大きく、竜脚形類、鳥盤類、テリジノサウルス類のような4本指の状態に近い。
著者らは4通りのデータセットを用いて分岐分析を行ったが、いずれもチレサウルスは獣脚類という結果になった。特に、テリジノサウルス類ではなくて、最も基盤的なテタヌラ類の位置にきた。
チレサウルスが獣脚類であることは、頸椎と前方の胴椎に側腔 pleurocoelがあること、胸部の脊椎骨にヒパポフィシスhypapophysisがあること、大腿骨の第4転子が半円形であることなどから支持されるという。チレサウルスがテタヌラ類であることは、肩甲骨が細長くひも状にのびていること、手根骨が半月形であること、手の第3指が縮小していることから支持される。
チレサウルスは、後肢とくに足首と足に、基盤的竜脚形類と似た驚くほど原始的な特徴をもっている。これは、走行にあまり適していない形態への二次的な逆戻りと考えられる。また、竜脚形類やテリジノサウルス類と共通した歯骨や歯の特徴は、植物食に関連した収斂と考えられる。恥骨が後方を向くことは、大量の植物を消化するための消化管の容積増大と関係する。
チレサウルスの発見によって、基盤的獣脚類における肉食から植物食への変化は、従来想像されたよりもよく起こる現象であったという考えが支持された。コエルロサウルス類以外では、植物食はこれまでリムサウルスしか知られていなかったが、チレサウルスが新たに植物食獣脚類のリストに加わることになった。
ニュースを見てファルカリウスみたいなものかと思っていたら、はるかに原始的だった。植物食であっても捕食者に狙われた場合、速く逃げられる方が有利な気もするが。オルニトミムス類のような方向性もあっただろう。走行に適さない形態への逆戻りということは、チレサウルスはあくまでテリジノサウルス系の方向性を目指したということか。テリジノサウルス類の場合は姿勢が変化しているので、後肢で体重を安定して支える必要があったが、チレサウルスも上体を起こして高い木の葉を食べたのだろうか。
参考文献
Fernando E. Novas, Leonardo Salgado, Manuel Suarez, Federico L. Agnolin, Martin D. Ezcurra, Nicolas R. Chimento, Rita de la Cruz, Marcelo P. Isasi, Alexander O. Vargas & David Rubilar-Rogers (2015) An enigmatic plant-eating theropod from the Late Jurassic period of Chile. doi:10.1038/nature14307
チレサウルスは、ジュラ紀後期チトン期(Toqui Formation)に南米のチリ南部に生息した基盤的テタヌラ類で、2015年にNovas et al.により記載された。植物食に適応した歯や腰帯、後肢などには、原竜脚類、テリジノサウルス類、はては原始的鳥盤類とも似た特徴がモザイク的に入り混じっているが、系統解析の結果から獣脚類とされた。ちなみに、一部ネットメディアの見出しのような「ティラノサウルスの近縁種」ではない。2本指であることはティラノサウルスとの類縁とは直接関係がない。前肢の指の数が減る傾向は獣脚類らしいといえばそうかもしれないが、もちろん主な根拠ではない。
チレサウルスのホロタイプ(SNGM-1935)は完全に近い、関節した全身骨格で全長約1.6 mと推定されるが、これはまだ成熟していない亜成体で、最も大きい標本の50%の大きさである。パラタイプは成長段階の異なる4個体の部分骨格で、全長1.2 から 3.2 mであるという。
チレサウルスに固有の特徴は、歯が葉状 leaf-shaped で萌出中の歯の歯冠の頂端にのみ鋸歯がある、烏口骨が四角形に近く厚い縁をもつ、手の第2指の末節骨の手前の指骨が短い、第3指が退化している、恥骨が完全に後方を向いている、恥骨の軸が棒状 rod-like で遠位端がふくらんでいない、などである。
頭骨は、分離した前上顎骨、上顎骨、前頭骨、後眼窩骨、歯骨などが見つかっており、頭は比較的小さかったと考えられる。前上顎骨は短く丈が高く、前端に粗面があり角質のくちばしがあったらしい。歯骨は前端の方が丈が高いので、テリジノサウルス類のように腹側縁が下方にカーブしている。歯骨の歯は丈が高く、葉状で、前方に傾いておりprocumbent、歯冠の頂端にのみ小さな鋸歯がある。
頸椎は長く、細長い首をなす。頸椎と前方の胴椎には、一対の仕切りのある側腔 pleurocoelがある。
四肢の骨は原竜脚類のようにがっしりしている。上腕骨は短く幅が広い。中手骨は第1から第3まであるが、よく発達しているのは第1指と第2指のみである。第1指の末節骨は他のテタヌラ類ほどカーブしていない。第2指の末節骨の手前の指骨が強く短縮している。厳密には2本指ではなく、第3指には中手骨だけではなく、1個の小さい指骨がある。(これは論文の写真には示されておらず、supplementary informationにもない。)
腸骨には竜脚形類、テリジノサウルス類、コエルロサウルス類、いくつかの鳥盤類と共通の特徴がある。恥骨は鳥盤類、テリジノサウルス類、ドロマエオサウルス類と同様に、完全に後方を向いている。恥骨の先端がふくらんでいないことは、多くの獣脚類とは対照的である。
足根部(足首)は基盤的竜盤類に似ている。距骨の上方突起は低く太く、板状で高く伸びているテタヌラ類とは異なる。ヘレラサウルスや基盤的竜脚形類のような初期の竜盤類と同様に、チレサウルスの踵骨は遠位側からみて三角形に近い。獣脚類の踵骨は長方形である。
チレサウルスの第1中足骨は第2中足骨の50%以上あり、これは他の多くの獣脚類と異なる。足の第1指は大きく、竜脚形類、鳥盤類、テリジノサウルス類のような4本指の状態に近い。
著者らは4通りのデータセットを用いて分岐分析を行ったが、いずれもチレサウルスは獣脚類という結果になった。特に、テリジノサウルス類ではなくて、最も基盤的なテタヌラ類の位置にきた。
チレサウルスが獣脚類であることは、頸椎と前方の胴椎に側腔 pleurocoelがあること、胸部の脊椎骨にヒパポフィシスhypapophysisがあること、大腿骨の第4転子が半円形であることなどから支持されるという。チレサウルスがテタヌラ類であることは、肩甲骨が細長くひも状にのびていること、手根骨が半月形であること、手の第3指が縮小していることから支持される。
チレサウルスは、後肢とくに足首と足に、基盤的竜脚形類と似た驚くほど原始的な特徴をもっている。これは、走行にあまり適していない形態への二次的な逆戻りと考えられる。また、竜脚形類やテリジノサウルス類と共通した歯骨や歯の特徴は、植物食に関連した収斂と考えられる。恥骨が後方を向くことは、大量の植物を消化するための消化管の容積増大と関係する。
チレサウルスの発見によって、基盤的獣脚類における肉食から植物食への変化は、従来想像されたよりもよく起こる現象であったという考えが支持された。コエルロサウルス類以外では、植物食はこれまでリムサウルスしか知られていなかったが、チレサウルスが新たに植物食獣脚類のリストに加わることになった。
ニュースを見てファルカリウスみたいなものかと思っていたら、はるかに原始的だった。植物食であっても捕食者に狙われた場合、速く逃げられる方が有利な気もするが。オルニトミムス類のような方向性もあっただろう。走行に適さない形態への逆戻りということは、チレサウルスはあくまでテリジノサウルス系の方向性を目指したということか。テリジノサウルス類の場合は姿勢が変化しているので、後肢で体重を安定して支える必要があったが、チレサウルスも上体を起こして高い木の葉を食べたのだろうか。
参考文献
Fernando E. Novas, Leonardo Salgado, Manuel Suarez, Federico L. Agnolin, Martin D. Ezcurra, Nicolas R. Chimento, Rita de la Cruz, Marcelo P. Isasi, Alexander O. Vargas & David Rubilar-Rogers (2015) An enigmatic plant-eating theropod from the Late Jurassic period of Chile. doi:10.1038/nature14307
コメント ( 2 ) | Trackback ( 0 )
モノロフォサウルスの再記載(頭骨)

少し大きい画像
{kind=link}
テタヌラ類の起源と初期の進化を探るには、ジュラ紀前期から中期が重要な時期であるが、この時期の化石記録は乏しい。その中で、中国のジュラ紀中期の獣脚類化石は非常に重要であるが、記載が不十分なために最近の系統研究には用いられないことが多い。
モノロフォサウルスは、ジュラ紀中期に中国新疆ウイグル自治区(ジュンガル盆地)に生息したテタヌラ類で、Zhao & Currie (1993) によって最初に記載された。Zhao & Currie (1993)は、モノロフォサウルスの形態には原始的な特徴と進歩的な特徴が入り混じっているとし、「アロサウルスに近縁なメガロサウルス段階‘megalosaur-grade’の獣脚類」とした。その後の多くの系統研究は、モノロフォサウルスをアロサウルス上科の中に位置づけてきた。しかし最近になって、Smith et al. (2007) の系統解析では、モノロフォサウルスはもっと基盤的なテタヌラ類とされた。そこでBrusatte et al. (2010) は、モノロフォサウルスの頭骨について、より詳細な再記載を行った(胴体については別の論文で記載)。
モノロフォサウルスの特徴は、一般向けには「頭骨の正中線上に一枚のとさかがある」で十分であるが、グァンロンなどが知られている現在、学問的にはどのように表現されるのだろうか。
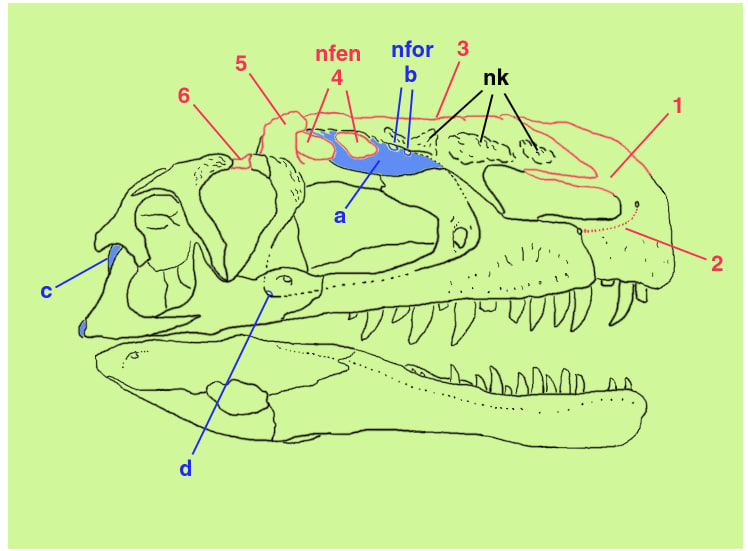
Brusatte et al. (2010) によると、モノロフォサウルスの頭骨についての固有の特徴は、(1)前上顎骨の鼻骨突起が、後方の鼻骨と接する部分で二つに分岐している、(2)前上顎骨の外側面に、鼻孔下孔subnarial foramenから鼻骨突起の基部の孔まで達する溝がある、 (3)鼻骨が盛り上がってとさかとなり、その背側縁はまっすぐで、上顎骨の腹側縁とほとんど平行である、 (4)鼻骨のとさかに2つの大きな、同じ大きさの含気窓pneumatic fenestraeがある、 (5)涙骨の背方に突出した、はっきりしたつまみ状の突起tab-like processがある、(6)左右の前頭骨を合わせると長方形で、長さよりも幅の方がずっと大きい(幅と長さの比率1.67)、であるという。
とさかの他に、外鼻孔が非常に大きいことも特徴的である。これについてはティラノサウルス上科のグァンロンとプロケラトサウルスも、よく似た大きい外鼻孔をもっている。
Brusatte et al. (2010) は今回新たに得られたデータを含めて、Smith et al. (2007) の解析をもとに修正した系統解析を行い、やはりモノロフォサウルスはアロサウルス上科ではなく、より原始的なテタヌラ類と結論している。
Smith et al. (2007) は、モノロフォサウルスをアロサウルス上科とする根拠として用いられてきたいくつかの形質は、実はアロサウルス上科以外の獣脚類にもみられることを指摘した。Brusatte et al. (2010) はさらに、アロサウルス上科の共有派生形質とされている多くの形質について、一つ一つ検討している。例えば、モノロフォサウルスではアロサウロイドと同様に、(a)前眼窩窩が鼻骨の側面に達している。しかしこの特徴は、原始的な獣脚類であるクリオロフォサウルス、ディロフォサウルス、マジュンガサウルスにもみられるという。またモノロフォサウルスとアロサウロイドでは(b)鼻骨の側面に含気孔pneumatoporeがある。しかし少なくともアベリサウルス類のマジュンガサウルスでは含気孔があり、また多くの基盤的獣脚類では鼻骨が見つかっていないため、この形質の共有派生形質としての使用は限定されるという。(c)方形骨が短いこともアロサウルス上科の共有派生形質とされており、モノロフォサウルスとアロサウロイドにはみられるが、他の獣脚類にもみられることがわかってきた。(d)頬骨の前眼窩窩の後腹側端に含気孔があることもアロサウルス上科の共有派生形質とされたことがあるが、これは大部分のアロサウロイドと基盤的なコエルロサウルス類であるティラノサウロイドにみられることから、アロサウルス上科の共有派生形質ではなく、より大きいグループ(アロサウルス上科+コエルロサウルス類)の共有派生形質とも考えられる。このように検討していくと、確実にアロサウルス上科の共有派生形質といえるものはほとんど残らず、アロサウルス上科が本当に単系統なのかという重大な問題になってくるという。
一方、モノロフォサウルスの頭骨の原始的な形質としてSmith et al. (2007) は、後眼窩骨が眼窩の下端に達していること、鼻骨・涙骨に加えて前上顎骨が参加するとさかがあること、方形骨と方形頬骨の縫合が側面に露出することを挙げた。Brusatte et al. (2010) はさらに、涙骨に含気性の兆候がないこと、前眼窩窩に副次的な孔が一つしかないこと、頭骨が長いこと(長さと高さの比が3.0に近い)などを挙げている。
あらためて側面図を見るとなかなかいい顔をしている。とさかには、鼻骨窓nasal fenestra (nfen), 鼻骨孔nasal foramen (nfor), 鼻骨こぶnasal knob (nk) などの部分があり、複雑な構造をしている。ディスプレイの他に上顎を強化する役割もあったのだろう。後の小型のティラノサウロイドのとさかを先取りするような進化をとげていた、といえるかもしれない。
参考文献
Brusatte, S. L., Benson, R. B. J., Currie, P. J., Zhao, X-J. (2010) The skull of Monolophosaurus jiangi (Dinosauria: Theropoda) and its implications for early theropod phylogeny and evolution. Zoological Journal of the Linnean Society, 158, 573-607.
コメント ( 0 ) | Trackback ( 0 )