獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2025年1月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | |||||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| 12 | 13 | 14 | 15 | 16 | 17 | 18 | ||
| 19 | 20 | 21 | 22 | 23 | 24 | 25 | ||
| 26 | 27 | 28 | 29 | 30 | 31 | |||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
アフロヴェナトル修正版
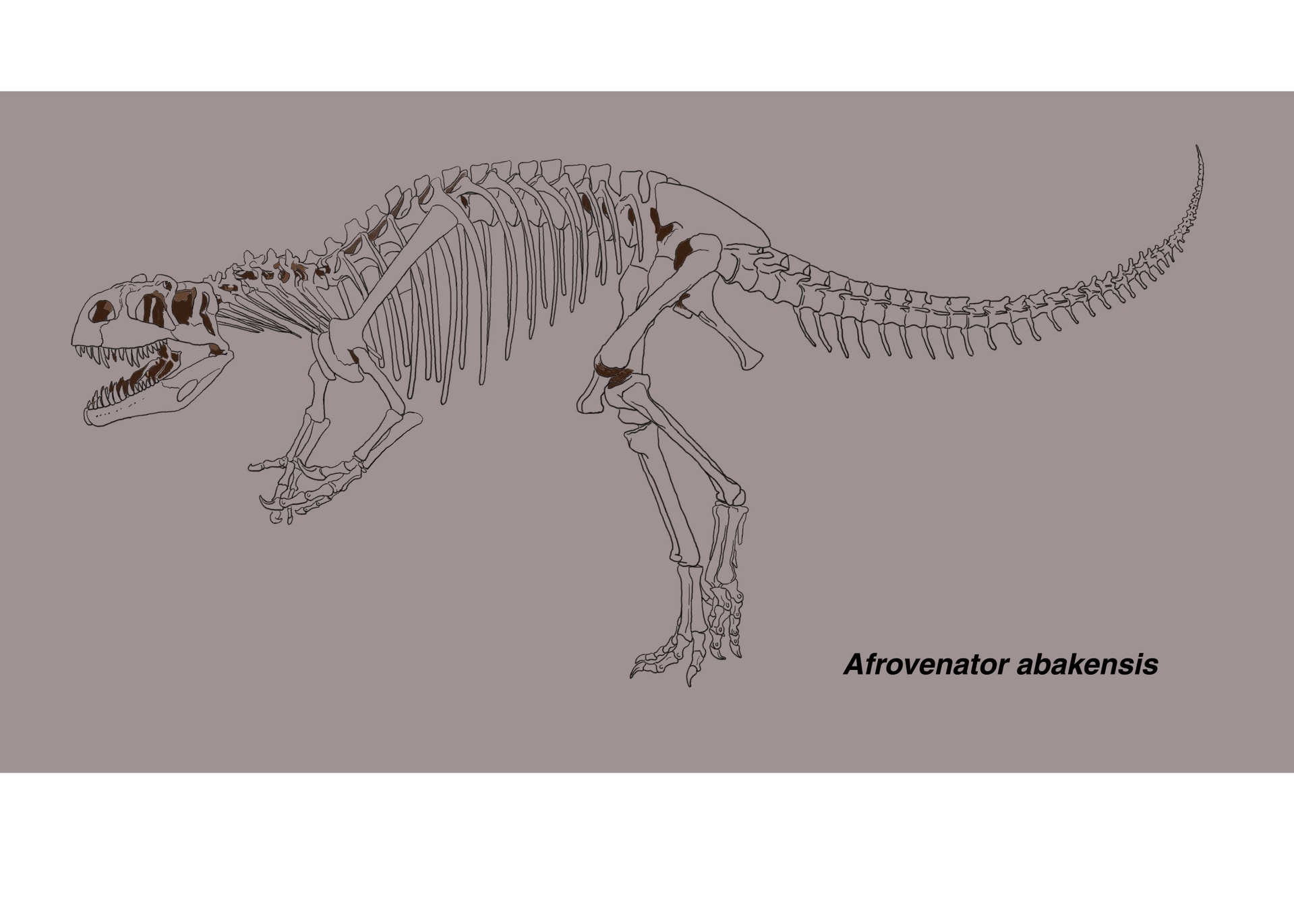
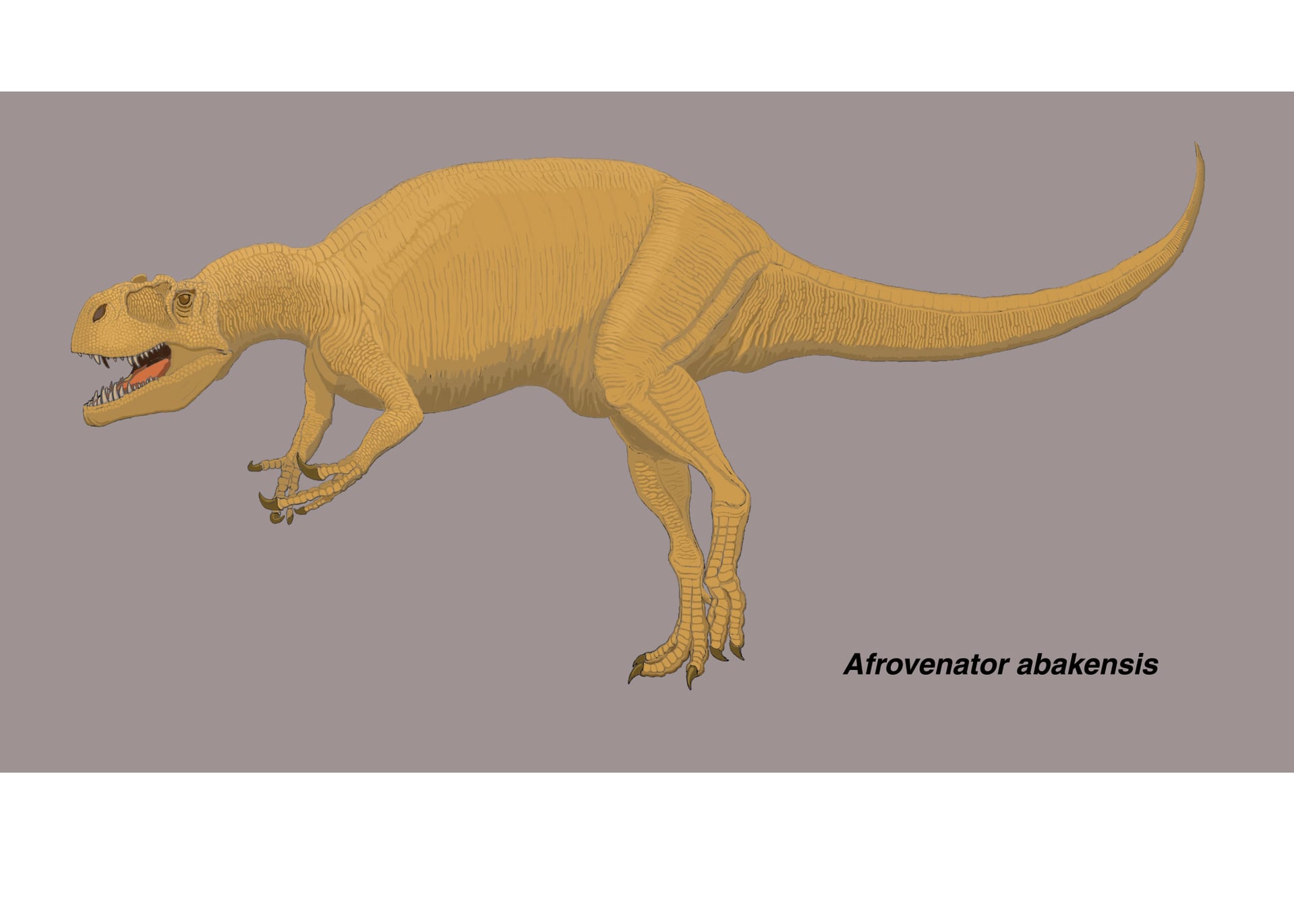
アフロヴェナトルも好きな恐竜の一つであるが、今年の科博の震災復興コラボミュージアムで、アフロヴェナトルが福島へ進出したようである。(日程の都合で行けなかった。)前にアロサウルスはやったということと、おそらく会場スペースの関係で手頃な大きさということか、肉食恐竜と植物食恐竜各1種ずつの枠に見事、選ばれた。大変喜ばしい。どうでもいいが今回「アフロベナトール」ではなくて「アフロヴェナトル」表記だった。福島の方々にもティラノやアロサウルスなどだけではなく、アフロヴェナトルの良さを(そもそも存在を)知っていただければ幸いである。メガロサウルス類で復元骨格が見られるのは、トルボサウルスとこのアフロヴェナトルくらいである。
コメント ( 0 ) | Trackback ( 0 )
トルボサウルスの最近の進展(2)アメリカ産とポルトガル産
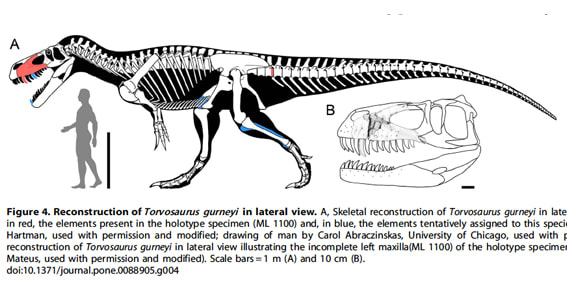
Copyright 2014 Hendrickx and Mateus
ポルトガルのロウリニャ層で発見された上顎骨と断片的な尾椎は、2006年にはアメリカのトルボサウルスと同種として報告されていた。その後、Hendrickx and Mateus (2014)がこの標本をアメリカのトルボサウルス・タンネリと詳細に比較した結果、いくつかの重要な違いが見出されたので、トルボサウルス属の新種トルボサウルス・グルネイとして記載された。
トルボサウルス・グルネイのホロタイプは、不完全な左の上顎骨と近位の尾椎の後方部分である。その他、別に見つかった脛骨と大腿骨の断片、分離した歯なども暫定的にトルボサウルス・グルネイと考えられている。
トルボサウルス・グルネイはほとんど上顎骨しかないので、著者らは世界中の大型獣脚類の上顎骨を比較研究している。多くの進化した獣脚類では、上顎骨の前眼窩窩にmaxillary fenestraという孔があいている。(わからない方は過去の記事の「アロサウルスの成長」「アリオラムスの頭骨を観察しよう」、「ヴェロキラプトル」、「丸ビルでアクロカント」などに図がある)トルボサウルスではこれが貫通した孔ではなく、浅い凹みになっている(「トルボサウルス1」参照)。
上顎骨の歯が収まっている歯槽の内壁は、歯間板 interdental plateという構造でできている。多数の歯間板が前後に並んでいるが、一部の獣脚類では歯間板が互いに癒合してひとつながりの板になっている。トルボサウルスでも歯間板が癒合して歯間壁interdental wallという構造になっている。
Hendrickx and Mateus (2014)はトルボサウルス属、トルボサウルス・タンネリ、トルボサウルス・グルネイの特徴を記述しているので簡単に記録する。
トルボサウルス(属)の特徴は、上顎骨の前眼窩窩に(貫通したmaxillary fenestraではなく)浅いmaxillary fossaをもつこと、前眼窩窩の前端腹側でmaxillary fossaの下に隆起した稜があること、歯間壁が上顎骨の内側面の1/2を占めること、などである。
トルボサウルス・タンネリの特徴は、歯間板の腹側縁が広いV字形で、歯間壁が上顎骨の外側壁より短い(上下にずれている)、内側棚の前方部で前内側突起anteromedial processの後方に隆起がある、である。
トルボサウルス・グルネイの特徴は、上顎骨の歯槽の数が11より少ない、歯間板の腹側縁がほぼ直線状、歯間壁が上顎骨の外側壁とほとんど一致している(揃っている)、内側棚の前方部で前内側突起anteromedial processの後方に隆起がない、である。(論文ではメガロサウロイドの中で固有の形質と、タンネリとの違いに分けて書いてあるが、重複するのでここではまとめて書く)
トルボサウルス・タンネリとトルボサウルス・グルネイの違いのうち、最もわかりやすいのは歯の数である。Britt (1991) によると、トルボサウルス・タンネリの上顎骨には11個の歯槽が観察され、全部で推定12-13個と考えられた。Hendrickx and Mateus (2014)の観察では確実なのは10個で、推定11-12個と考えている。それに対してトルボサウルス・グルネイでは8個しかみられず、最大でも10個と考えられた。歯の数は同種内でも個体によって変異がみられるので、これを種の特徴とするのには慎重でなければならないが、メガロサウロイドの中で上顎骨の歯の数が11より少ないのはトルボサウルス・グルネイだけである。
トルボサウルス・タンネリでは、歯間板の腹側縁が広いV字形をしている。一方トルボサウルス・グルネイでは歯間板の腹側縁が直線状で、歯間壁全体にわたって隣と連続している。V字形の歯間板は、多くの獣脚類にふつうにみられる。例えばノアサウルス類マシアカサウルス、アベリサウルス類ルゴプス、多くのメガロサウルス類、アロサウロイド(アロサウルス、ネオヴェナトル、シンラプトル、マプサウルス)、ティラノサウルス類(アリオラムス、タルボサウルス、ティラノサウルス)などである。一方、直線状(長方形)の歯間板は、ケラトサウルス類(ケラトサウルス、ノアサウルス、アウカサウルス、マジュンガサウルス)、メガロサウルス類メガロサウルス、アロサウロイドのシャオチーロン、ティラノサウロイドのエオティランヌスにみられる。
もう一つの違いは上顎骨の外側縁に対する、歯間板(の癒合した歯間壁)の腹側への伸展の度合いである。トルボサウルス・グルネイでは、歯間壁は上顎骨の外側縁とほとんど同じくらいまで伸びている。一方トルボサウルス・タンネリでは、歯間壁は短く、上顎骨の外側縁よりもかなり上(背側)で終わっている。このような歯間板の相対的な伸展の程度は、ケラトサウルス、マジュンガサウルス、メガロサウルス、アロサウルスなどの同種内の異なる標本の間では違いはみられなかった。それに対してカルカロドントサウルス・サハリクスとカルカロドントサウルス・イグイデンシスの間では違いがあり、前者の歯間板の方がより腹側まで伸びている。このことから歯間板の相対的な伸展は種間で異なるものであり、2種のトルボサウルスを識別する特徴と考えられるという。


大きい画像
参考文献
Hendrickx C, Mateus O (2014) Torvosaurus gurneyi n. sp., the Largest Terrestrial Predator from Europe, and a Proposed Terminology of the Maxilla Anatomy in Nonavian Theropods. PLoS ONE 9(3): e88905. doi:10.1371/journal.pone.0088905
コメント ( 0 ) | Trackback ( 0 )
トルボサウルスの最近の進展(1)博物館に眠っていた化石
トルボサウルスの四肢に関する情報としては、Hanson and Makovicky (2014) が出版されている。しかし、この論文はトルボサウルスの完全な前肢が関節状態で見つかった、というような話ではない。むしろ断片的なものである。
これは、100年以上も前に発掘された化石の再発見である。この化石標本は1899年にElmer Riggsの調査隊によって、ワイオミング州のFreezeout Hillsで発見されたが、その後手つかずのまま、フィールド自然史博物館の収蔵庫に保管されていた。在庫目録の作成の際に、重要な化石かもしれないということで、2005年にクリーニングされた。
この標本FMNH PR 3060は、3つの腹肋骨(ガストラリア)の断片、右の第III中手骨、右の指骨III-2、左の中足骨II, III, IV, 左の趾骨I-1からなる。化石の発見された地層は、モリソン層のBrushy Basin Member(ジュラ紀後期チトン期初期、150-148 Ma)に属し、これまで知られているトルボサウルスの生息時期と一致する。
この標本は、中足骨の大きさとプロポーションに基づいて、モリソン層の獣脚類の中でもトルボサウルス・タンネリと同定できる。トルボサウルスでは中足骨の長さが近位端の幅の2~3倍であるが、アロサウルスとケラトサウルスでは3~4倍である。またcollateral ligament pitが広く浅いことは、トルボサウルス、エウストレプトスポンディルス、ピアトニツキサウルスのようなメガロサウルス類にみられる形質であり、アロサウルスやケラトサウルスとは異なる。
これらのうち、後肢の趾骨I-1と前肢の指骨III-2はトルボサウルスでは初めて見つかった骨である。前肢の指骨III-2が知られている他のメガロサウルス類は、スキウルミムスしかいない。この指骨は、骨幹が短縮して三角形に近い形をした、最も短い指骨である点でスキウルミムスの同じ骨と非常によく似ている。
この標本には前肢の手の骨と後肢の足の骨が含まれているが、メガロサウルス類の手・足の標本は非常に乏しいために、手と足の比率などを他のメガロサウルス類と比較するのは困難である。手と足の両方の骨が十分保存されているメガロサウルス類は、幼体であるスキウルミムスだけである。Rauhut et al. (2012) によると、スキウルミムスのプロポーションはジュラヴェナトルのようなコエルロサウルス類の幼体と驚くほど似ているという。その特徴の一つとして、比較的手が長いことがあげられる。スキウルミムスでは手の第2指の長さ(中手骨を含み末節骨は含まない)が第3中足骨の長さと同じくらいである。しかしこの比率はメガロサウルス類の成体には当てはまらないだろうという。Britt (1991) は、トルボサウルスの中手骨や他の前肢の骨は、獣脚類としては比較的短く太いといっている。
今回の標本は、これまでに記載されたトルボサウルスの標本と同じくらいの大きさである。Jensen (1985)によって記載された標本の第3中足骨の長さは320-365 mmであるが、FMNH PR 3060では356 mmで、この範囲に入っている。今回の標本を含めて、これまでモリソン層で見つかったトルボサウルスの標本はみな同じくらいの大きさの成体であるという。
比較できる足の骨が知られているメガロサウルス類としてはエウストレプトスポンディルス、メガロサウルス、アフロヴェナトル、ポエキロプレウロン、スキウルミムスなどがあるが、トルボサウルスの足の骨はこれらのどれよりもがっしりしている。比較できる手の骨が知られているメガロサウロイドはスコミムス、レシャンサウルス、スキウルミムスに限られる。スコミムスの第3中手骨はまだ記載されていない。レシャンサウルスとスキウルミムスの第3中手骨は、トルボサウルスよりも華奢なものである。
参考文献
Michael Hanson & Peter J. Makovicky (2014). A new specimen of Torvosaurus tanneri originally collected by Elmer Riggs. Historical Biology: An International Journal of Paleobiology, 26:6, 775-784, DOI: 10.1080/08912963.2013.853056
コメント ( 0 ) | Trackback ( 0 )
トルボサウルスの卵殻と胚:恐竜の卵殻形態の系統的ギャップが埋められる
これまで非鳥型竜盤類で、卵殻と胚が一緒に見つかっているのは、ほとんど竜脚形類マッソスポンディルスとコエルロサウルス類に限られていて、基盤的獣脚類の知見がなかった。Arau´jo et al. (2013) はポルトガルのロウリンニャ層から、メガロサウルス類トルボサウルスと考えられる胚の骨格と卵の化石を発見し、Scientific Reportsに報告している。
卵と胚が共に化石として発見されることは非常にまれであり、竜盤類では1)前期ジュラ紀の竜脚形類マッソスポンディルス、2)後期ジュラ紀の獣脚類ロウリンハノサウルス、3)後期白亜紀のテリジノサウルス類、4)前期および後期白亜紀のティタノサウルス類、5)後期白亜紀のトロオドン類、6)後期白亜紀のオヴィラプトル類、で報告されている。すなわち、基盤的な獣脚類など竜盤類の根元あたりの情報がごっそり抜けている。
著者らはシンクロトロン放射式マイクロCT、走査型電子顕微鏡、光学顕微鏡などを駆使して、卵殻の形態と特定のグループの基盤的獣脚類の骨学形態を結びつけている。
標本は直径65 cmの集合体で、500以上の卵殻の破片と胚の骨と歯からなっている。2005年にロウリンニャ層Lourinha˜ FormationのSobral Memberから発見され、2005から2006年に発掘され、2009年にロウリンニャ博物館でクリーニングされた。標本には方向の異なる3つの塊が含まれ、いずれもひどく割れているが、3つ以上の卵の集まりと考えられた。胚の骨としては5本の分離した歯、4本の歯のついた上顎骨、4本の歯のついた歯骨、1本の分離した歯骨歯、3個の関節した椎体、その他の同定できない骨が含まれていた。
どうしてこれがトルボサウルスとわかるのだろうか。この標本はまず、非常に長く、先端がカーブして鋭く尖った歯冠をもつことから獣脚類と考えられる(鋸歯はみられなかった)。歯列が眼窩より前方に限られることからテタヌラ類であることが示唆される。また上顎骨の頬骨突起の背側縁と腹側縁が急速にすぼまっていることなどから、メガロサウルス科と考えられた。さらに歯骨の歯の数(下顎結合の前端からMeckelian fossaの前端までの)が10より少ないことなどから、メガロサウルス亜科Megalosaurinaeと思われた。
多くのテタヌラ類では上顎骨を貫通するmaxillary fenestraがあるが、メガロサウルスやドゥリアヴェナトルでは上顎骨の内側の小さい孔となっている。しかしこの標本のように、窓も含気窩もない上顎骨はメガロサウルス科の中でも非常に限られている。
この胚は、1)上方突起の基部より後方に(maxillary fenestraなどの)含気構造がない、2)上方突起の基部と上顎骨の腹側縁の間の角度が35°より小さい、3)上顎骨の頬骨突起の先端が舌状の形をしていることから、トルボサウルス属と考えられた。これらの特徴はメガロサウルスにはあてはまらない。さらにトルボサウルスは、以前ポルトガルの同じロウリンニャ層から発見されている。
ただし、この胚にはトルボサウルスの成体とは異なる点が4つある。1)歯の前縁にも後縁にも鋸歯がない、2)上顎骨の歯間板が癒合していない、3)上顎骨の前方突起が短い、4)上顎骨の歯が6より少ない、である。これらの違いは個体発生上の成長段階による形態変異と考えられた。
卵殻の厚さは約1.2 mmで、次のような特徴がある。(1)表面に、血管網のように互いに吻合した、網目状の装飾構造anastomizing ornamentationがある、(2)卵殻の外側に向かって放射状に並んだ、針状ないし長い剣状の炭酸カルシウムの結晶(方解石)がある、(3)ほぼ1層しかない。多くの場合、卵殻の基底層にmammillae乳頭状突起というものがあるらしいが、この卵殻でははっきりしないらしい。
卵の化石にはそれなりの形態分類があるようで、今回の卵殻は、Dendroolithidae oofamily の卵に最もよく似ている。Dendroolithidae の卵殻は従来、竜脚類か鳥脚類のものと考えられていたが、今回の胚の形態は明らかにこのタイプの卵殻を獣脚類と結びつけるものである。
系統発生学的意義については次のように述べている。まず鳥類(アヴィアラエ)の卵殻は3層構造をしている。コエルロサウルス類(しかもマニラプトル類)であるトロオドン類とオヴィラプトル類の卵殻は2層構造である。一方、竜脚類ティタノサウルス類の卵殻は1層である。今回のメガロサウルス類トルボサウルスの卵殻は、獣脚類で初めて報告された1層の卵殻であり、これが獣脚類の原始状態plesiomorphic conditionと考えられる。また1層構造と針状の形態は竜盤類の共有派生形質と思われる。
そこで問題となるのが、同じロウリンニャで発見されている暫定的にロウリンハノサウルスの卵とされている化石である。ロウリンハノサウルスもアロサウロイドなどの基盤的テタヌラ類とされているが、卵殻の構造が全く異なっている。ロウリンハノサウルスの卵殻は表面の装飾構造がなく、コエルロサウルス類のような2層構造である。このことから1)これがロウリンハノサウルスの卵殻であるという同定が間違っている、2)テタヌラ類では卵殻の構造が非常に変化しやすい、3)ロウリンハノサウルスの系統的位置が不明確である(実はコエルロサウルス類である可能性など)、ことが示唆される。
最後に余談であるが、このように上顎骨にmaxillary fenestra やpromaxillary fenestra などの穴がないことが、トルボサウルスの重要な特徴である。同定の根拠とされるほど重要な特徴である。もちろん2006年に報告されたポルトガルのトルボサウルス(ジュラ紀最大の獣脚類という)の上顎骨にも穴はない。
ところで、レプリカ販売会社 Zoic srl のウェブサイトhttp://www.zoic.it/zoic/zoic.html にはTorvosaurus sp. の復元頭骨がある。これはコロラド州ドライ・メサの新しい標本らしいが、上顎骨に2つ穴がある。しかも貫通しているような…??これは一体どういうことですかね。単にいいかげんな作りとすれば、トルボサウルスに失礼である。今度、バッキア氏に訊いてみようか。
参考文献
Ricardo Araujo, Rui Castanhinha, Rui M.S. Martins, Octavio Mateus, Christophe Hendrickx, F. Beckmann, N. Schell, & L.C. Alves (2013)
Filling the gaps of dinosaur eggshell phylogeny: Late Jurassic Theropod clutch with embryos from Portugal. SCIENTIFIC REPORTS 3 : 1924 / DOI: 10.1038/srep01924
Mateus, O., Walen, A. & Antunes, M. T. (2006) The large theropod fauna of the Lourinha˜ Formation (Portugal) and its similarity to the Morrison Formation, with a description of a new species of Allosaurus. N. Mex. Mus. Nat. Hist. Sci. Bull. 36, 123-129.
コメント ( 0 ) | Trackback ( 0 )
スキウルミムス

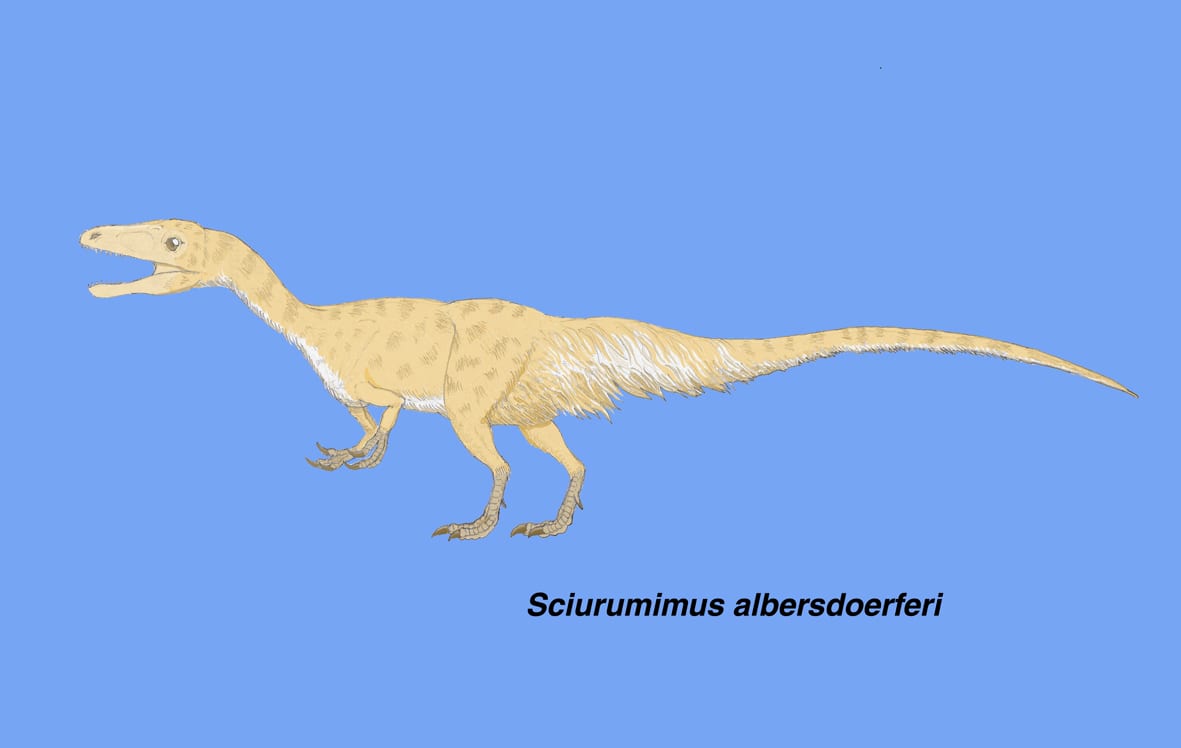
少し大きい画像
これまで羽毛の痕跡が見つかった獣脚類は、ほとんどが中国遼寧省のものであり、すべてコエルロサウルス類であった。
スキウルミムスは、ジュラ紀後期キンメリッジ期にドイツ南部バイエルン州に生息した獣脚類で、2012年に記載された。化石は保存の良い全身骨格で、孵化後まもない幼体と思われるが、体の各部に羽毛の痕跡が認められた。系統解析の結果、この恐竜はメガロサウルス類と考えられたことから、羽毛の起源がコエルロサウルス類よりもはるかに原始的な段階まで、さかのぼることが示唆された。
尾に長めの羽毛があることからスキウルミムス(リスもどき)と命名されたが、全体としてはリスには似ていない。仔ワニかトカゲというところである。
系統解析の結果、スキウルミムスは基盤的テタヌラ類であり、基盤的テタヌラ類の中の系統関係を詳細に解析すると、メガロサウルス上科の中でメガロサウルス科の基盤的なメンバーとなった。スキウルミムスにみられるメガロサウルス上科の共有派生形質は、上顎骨の前方突起が長い、maxillary fenestraが内側で閉じている、涙骨の前方突起が非常に細い、涙骨のlateral bladeが前眼窩窓の上に張り出していない、基底後頭顆の腹側に深い凹みがある、歯骨の前端がわずかに背方に膨らんでいる、前方の胴椎の腹側に顕著なキールがある、前肢の第1指の末節骨が拡大している、などである。
興味深いことに、従来のデータにスキウルミムスを加えた今回の解析結果では、単系の「カルノサウルス類」が復活している。つまりテタヌラ類がまず「カルノサウルス類」とコエルロサウルス類に分岐し、「カルノサウルス類」の中にメガロサウルス上科とアロサウルス上科が含まれる形になっている。この分岐図は、最近のほとんどの系統解析とは異なっており、スキウルミムスが幼体であることを考えると、慎重に考えなければならないと著者自身が述べている。いずれにしても1つの新種を加えるだけで、系統関係に重大な変更が起きてしまうということは、テタヌラ類の系統進化について我々がまだ十分理解していないことを示している、としている。
スキウルミムスは、最も完全に保存されたメガロサウルス類の全身骨格ということになり、これまで知られていなかったメガロサウルス類の解剖学的詳細を解明するのに役立つ。例えばスキウルミムスの手は3本指で、第4中手骨の痕跡はなかった。テタヌラ類は3本指であるが、原始的なアロサウロイドであるシンラプトル、ネオヴェナトル類メガラプトル、原始的なティラノサウロイドであるグァンロンでは、痕跡的な第4中手骨がみられる。このことから、第4中手骨の消失はいくつかの系統で独立して起こったか、あるいはテタヌラ類の祖先で消失したが、いくつかの種類で「先祖返り」的に第4中手骨が出現したと考えられる。
スキウルミムスの模式標本は、いくつかの特徴から非常に若い、おそらく孵化後まもない幼体と考えられた。例えば、頭骨が非常に大きく後肢が短いという体のプロポーションや、骨格に癒合がみられないこと(脊椎全体で神経椎体縫合が閉じていない、仙椎の癒合がない、脳函の各要素が癒合していない)、粗い溝のある骨表面形状、上顎骨の歯の成長パターンが非常に規則的であること(まだ生え替わっていないことを示す)などである。
スキウルミムスの歯の形態は、ほっそりした鋸歯のない前上顎歯と、後縁のみに鋸歯のある強く反った上顎歯をもつ点で、基盤的テタヌラ類の成体とは著しく異なる。このような特徴はおそらく幼体の形質と思われる。これは大型獣脚類の幼体は成体とは異なる獲物を食べていたという考えを支持する。むしろ、このような歯の形態は、基盤的なコエルロサウルス類と非常によく似ている。よって、このような形態の歯が見つかった場合にコエルロサウルス類と同定するには注意が必要と考えられる。
羽毛は尾、背中、腹部など数カ所に、皮膚の痕跡とともに保存されている。最も顕著なのは前方の尾椎の背側で、そこには非常に細長い、1型羽毛 type 1 feather に該当する毛状の繊維構造がある。この原羽毛は皮膚から生えているように見え、尾の背側で厚い被覆をなしている。また枝分かれしておらず、シノサウロプテリクスの原羽毛と同じくらいのサイズである。
羽毛の分布から、スキウルミムスはコンプソグナトゥス類と同様に、全身が羽毛で覆われていたと考えられる。スキウルミムスは、これまでに羽毛が報告された最も基盤的な獣脚類であり、少なくとも基盤的テタヌラ類の幼体には原羽毛があったことを示した。これは、繊維状構造が知られている鳥盤類(プシッタコサウルスやティアニュロン)とコエルロサウルス類の間のギャップを埋めるものである。鳥盤類の繊維も獣脚類の原羽毛と形態学的に区別できないことを考えると、羽毛をもつことは恐竜全体の原始形質と考えられる、という。成体でウロコの痕跡が見つかることは羽毛の存在と矛盾するものではなく、大型恐竜の成体では二次的に羽毛を失うこともあっただろう、としている。
参考文献
Oliver W. M. Rauhut, Christian Foth, Helmut Tischlinger, and Mark A. Norell (2012) Exceptionally preserved juvenile megalosauroid theropod dinosaur with filamentous integument from the Late Jurassic of Germany. PNAS 109, (29) 11746-11751; published ahead of print July 2, 2012, doi:10.1073/pnas.1203238109
コメント ( 0 ) | Trackback ( 0 )
アフロヴェナトル1
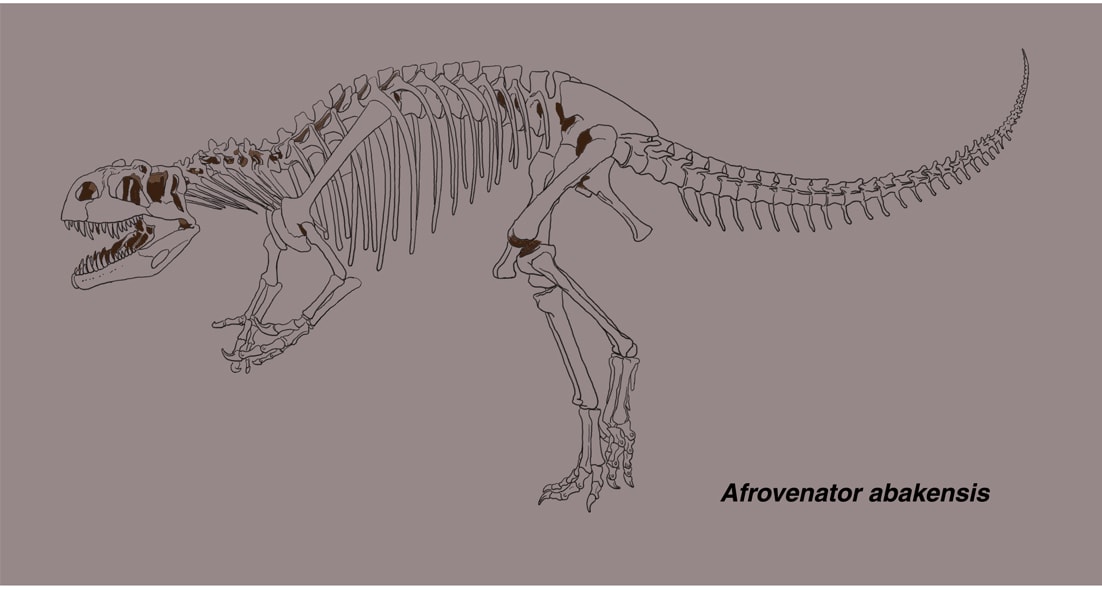
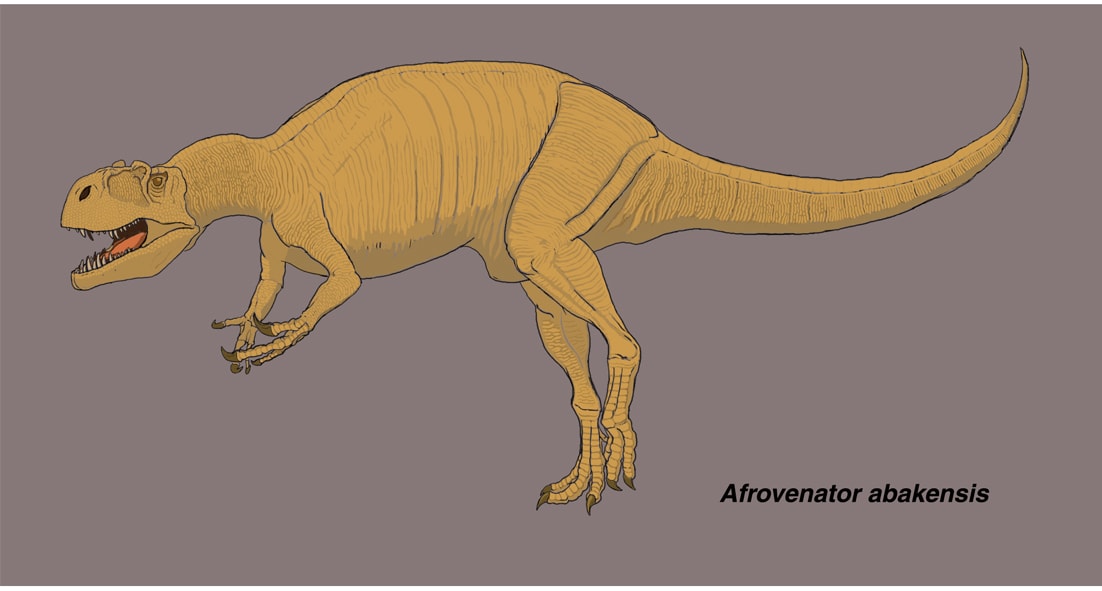
アフロヴェナトルは、ジュラ紀中期(バス期からオックスフォード期)に北アフリカのニジェールに生息したメガロサウルス類で、1994年に記載された。部分的な頭骨と体の骨格が知られている。全体としてアロサウルスと似ているが、個々の骨が細長く、より華奢なつくりとなっている。
アフロヴェナトルの固有の形質は、前眼窩窩の前縁が耳たぶ状になっている、第3頸椎の神経棘が低い長方形、非常に扁平な半月形の手根骨、第1中手骨の第2中手骨との関節面に広い突縁があることであるという。
前上顎骨と鼻骨は見つかっていないので推定であるが,頭骨は長く,丈が低い(長さに対する眼窩の位置での高さの比率が1/3以下)。他のメガロサウルス類でははっきりした涙骨角はないが、アフロヴェナトルには低い涙骨角があり、含気孔がある。上顎骨にはスリット状のpromaxillary fenestra と小さなmaxillary fenestraがあり、14本のナイフ状の歯がある。下顎はほとんど発見されていない。前肢はアロサウルスと同じくらいの長さと推定されているが、上腕骨が比較的長く、手根部がより扁平で、第3指の末節骨が小さい。撓骨と尺骨はほとんど見つかっていない(論文の骨格図では遠位端のみがあるように描かれているが、言及はない)。後肢はアロサウルスと比べて遠位の骨要素が長い。大腿骨に対する脛骨の比率は、アロサウルスの0.81に対してアフロヴェナトルでは0.90である。足(中足骨と趾骨)もアロサウルスより細長い。
1994年のセレノの記載では,前眼窩窩にmaxillary fenestraがある、上顎骨の歯列が眼窩の前方に限られるなどの形質からテタヌラ類であり、上顎骨の前方突起が長い、後眼窩骨の腹方突起が横に幅広いなどの形質からスピノサウルス上科(原文:トルボサウルス上科)とされた。前肢の第1指の末節骨がそれほど大きくないことから基盤的な種類として、メガロサウルス科(原文:トルボサウルス科)やスピノサウルス科の外に置かれた。
Allain (2002)は頭骨の45の形質に基づいて、メガロサウルス類を中心に基盤的なテタヌラ類の系統解析を行い、その結果、トルボサウルス、エウストレプトスポンディルス、ドゥブレウイロサウルス、アフロヴェナトルをメガロサウルス科としている。ここではスピノサウルス上科の共有派生形質は、後眼窩骨の腹方突起が眼窩の下縁近くまで延びていること、上顎骨の前方突起が長方形であること、頭骨の長さに対する眼窩の位置での高さの比率が1/3以下、などとしている。メガロサウルス科の共有派生形質は、後眼窩骨の腹方突起の断面がU字型であること、方形骨に孔がないこと、などとしている。さらにAllain (2002)ではメガロサウルス科をトルボサウルス亜科(トルボサウルスとおそらくエドマルカ)とメガロサウルス亜科(エウストレプトスポンディルス、ドゥブレウイロサウルス、アフロヴェナトル)に分けている。両者の違いは、前眼窩窩のpromaxillary recess (fenestra)の部分がトルボサウルスでは浅い窪みであるが、後の3種類では深い孔である(ただし内部で閉じている)ことなどである。(ここはアロサウルス、シンラプトルやコエルロサウルス類では内側まで貫通している。)
The Dinosauria 2nd ed. (2004) ではメガロサウルス科のうちエウストレプトスポンディルス亜科としてエウストレプトスポンディルス、ドゥブレウイロサウルス、アフロヴェナトル、ピアトニツキサウルスを含めている。共有する形質はmaxillary fenestraがあることなどとしている。
初めて見たのは1998年の科博(ゴンドワナ)で、イラストは2002年の恐竜博の全身復元骨格を元にしている。ところでアフロヴェナトルの前腕(撓骨・尺骨)はほとんど発見されていないようである。スピノサウルス上科では上腕骨に対して前腕が短いのが特徴とされている。現在メガロサウルス科の中に入っているのであれば、もう少し前腕が短くてもいいような気もする。あるいは前腕が短いのは、スピノサウルス科とトルボサウルスなどで独立して獲得した形質なのだろうか。
参考文献
Sereno, P. C., J. A. Wilson, H. C. E. Larsson, D. B. Dutheil, and H. D. Sues. 1994. Early Cretaceous dinosaurs from the Sahara. Science 266: 267-271.
Allain, R. 2002. Discovery of megalosaur (Dinosauria, Theropoda) in the Middle Bathonian of Normandy (France) and its implications for the phylogeny of basal Tetanurae. Journal of Vertebrate Paleontology 22:548-563.
コメント ( 2 ) | Trackback ( 0 )
トルボサウルス1続き
ドライ・メサからは頭骨のほか、頸椎、胴椎、尾椎、血道弓、前肢の骨(上腕骨、撓骨、尺骨、第1、2、3中手骨、第1指の第1指骨)、後肢の骨(腸骨、恥骨、座骨、脛骨、腓骨、距骨、踵骨、中足骨)が発見されている。しかしBritt (1991)のAPPENDIXのトルボサウルスの骨のリストには、なぜか上顎骨が抜けている。また、ホロタイプの上腕骨、撓骨、尺骨は含まれているのに、中手骨BYUVP2010-2012と指骨BYUVP2018は載っていない。さらに腰帯(腸骨、恥骨、座骨)も載っていない。この論文で記載している上顎骨や腰帯がリストから抜けているのは奇妙であるが、ミスなのだろうか。
肋骨、肩甲骨、烏口骨、大腿骨は見つかっていない。また前肢の指骨のほとんどと後肢の趾骨も見つかっていない。さらに、有名な大きな末節骨はBritt (1991)には記載されていないしリストにもない(第1指の末節骨は第2指骨のはず)。別に報告されているということか。
またGalton and Jensen (1979)ではホロタイプの上腕骨、撓骨、尺骨は同一個体のものと考えていたが、すべて関節状態ではなく、ドライ・メサからは3頭のトルボサウルスが出ていることを考えると、複数の個体が混じっている可能性もあるという。
北アメリカのモリソン層と似た動物相を産するポルトガルのロウリンニャ層では、2003年にプライア・ダ・ヴェルメリャから大型獣脚類の上顎骨が発見され、2006年に報告された。この上顎骨は、前眼窩窩に穴がなく、鼻骨突起が含気性でなく、鼻骨突起がやや後方に位置するなどの形質からトルボサウルスと同定された。上顎骨はがっしりしていて、8個の歯槽が保存されており、さらに2個の歯槽の存在が推定されるので、合計10本の歯があったと考えられる。コロラドのトルボサウルスは推定13本なので(11本なのかもしれない)数は異なるが、ポルトガル産の方も現在のところ同一種とされている。このポルトガル産の上顎骨は長さが63 cmもあり、頭骨は158 cmと推定された。もしそのとおりならばジュラ紀最大の獣脚類となるという。
参考文献
Britt, B. B. 1991. Theropods of Dry Mesa Quarry (Morrison Formation, Late Jurassic), Colorado, with emphasis on the osteology of Torvosaurus tanneri. Brigham Young University Geology Studies 37: 1-72.
DinoData.
肋骨、肩甲骨、烏口骨、大腿骨は見つかっていない。また前肢の指骨のほとんどと後肢の趾骨も見つかっていない。さらに、有名な大きな末節骨はBritt (1991)には記載されていないしリストにもない(第1指の末節骨は第2指骨のはず)。別に報告されているということか。
またGalton and Jensen (1979)ではホロタイプの上腕骨、撓骨、尺骨は同一個体のものと考えていたが、すべて関節状態ではなく、ドライ・メサからは3頭のトルボサウルスが出ていることを考えると、複数の個体が混じっている可能性もあるという。
北アメリカのモリソン層と似た動物相を産するポルトガルのロウリンニャ層では、2003年にプライア・ダ・ヴェルメリャから大型獣脚類の上顎骨が発見され、2006年に報告された。この上顎骨は、前眼窩窩に穴がなく、鼻骨突起が含気性でなく、鼻骨突起がやや後方に位置するなどの形質からトルボサウルスと同定された。上顎骨はがっしりしていて、8個の歯槽が保存されており、さらに2個の歯槽の存在が推定されるので、合計10本の歯があったと考えられる。コロラドのトルボサウルスは推定13本なので(11本なのかもしれない)数は異なるが、ポルトガル産の方も現在のところ同一種とされている。このポルトガル産の上顎骨は長さが63 cmもあり、頭骨は158 cmと推定された。もしそのとおりならばジュラ紀最大の獣脚類となるという。
参考文献
Britt, B. B. 1991. Theropods of Dry Mesa Quarry (Morrison Formation, Late Jurassic), Colorado, with emphasis on the osteology of Torvosaurus tanneri. Brigham Young University Geology Studies 37: 1-72.
DinoData.
コメント ( 1 ) | Trackback ( 0 )
トルボサウルス1
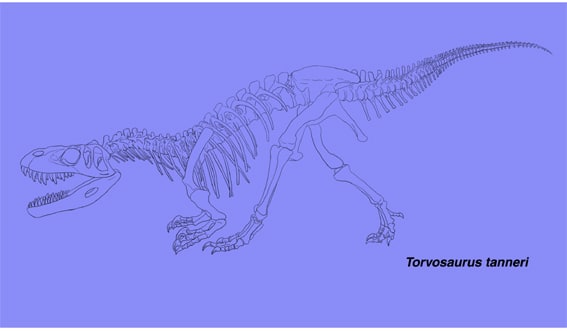
多くの恐竜ファンにとって、展示されている復元骨格のうち、どの部分が実際に発見されていて、どの部分が推定なのかということは、基本的に重要な関心事であると思うが、明確に示されていることは少ない。実際に見つかっている部分が少なく、大部分が近縁の種類に基づく推定である場合、「なあんだ、ほとんど作り物か」とがっかりされる(夢をこわす?)のを恐れて説明しないのだろうか。もしそうだとしたら、それは実は逆効果ではないだろうか。明らかに推定を含んでいる全身骨格に、何の説明もされていないと、実際に見つかっている部分でさえ信用できなくなってしまう。あるいは逆に、推定で作られた部分を、その種類の特徴と信じて一所懸命にスケッチする人もいるだろう。骨格図に色分けでもして示されていれば、少なくともこの部分は信頼できるんだな、と納得できるはずである。例えば四川省のスゼチュアノサウルスなども、昔はぎこちない感じのキャストだったが、模型制作技術が向上したのか、最近は妙にきれいな全身復元骨格が展示されるようになった。中型の獣脚類であるという以外に、どこをどのように信頼していいのか、かえってわからない。
トルボサウルスは、日本では2002年の恐竜博で初めて全身復元骨格が展示された。このときネット上には大変美しい、すばらしいという声とともに、どうも作り物っぽいという意見もあった。私も、下顎の形状などがのっぺりしていて、なにか「木製家具」の部品のような印象があって気になった。恐竜博の図録では一部の骨がイラストに示されているが、頭骨が(本文に特徴が書いてあるのに)示されていない。研究史を通じてどのくらいの骨が発見されているのだろうか。
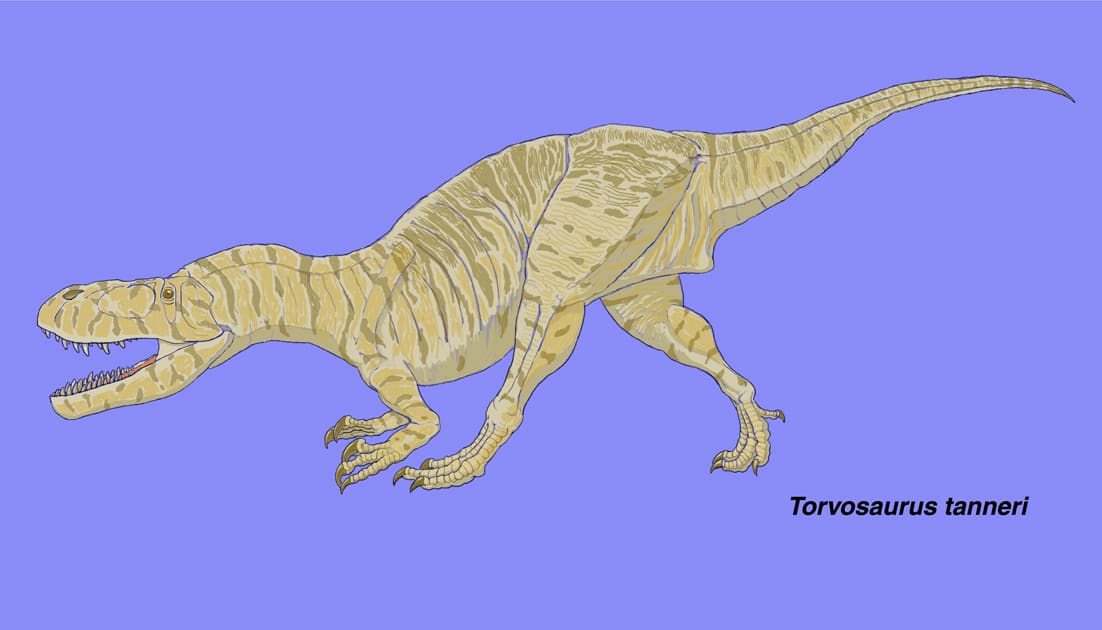
トルボサウルスは、ジュラ紀後期に北アメリカ(コロラド、ユタ、ワイオミングの各州)とヨーロッパ(ポルトガル)に生息したメガロサウルス類で、コロラド州ドライ・メサ採掘場から発見され1979年に記載された。ブリガムヤング大学のチームによって1972年から12期にわたり発掘が行われたが、トルボサウルスの化石はほとんどが第1期に採集された。関節していない状態で、区別できない同じ大きさの大型の2個体と、やや小さい1個体の3頭分の骨格が含まれていた。Galton and Jensen (1979)により完模式標本とされたのは前肢の上腕骨、撓骨、尺骨で、撓骨・尺骨の長さが上腕骨の約1/2しかないことは特徴の一つである。
トルボサウルスは、全長9メートルに達する大型でがっしりしたメガロサウルス類である。頭骨は中程度に長く、丈の高さが長さの約40%と推定されている。吻の先端は尖っている。前上顎骨に3本、上顎骨におそらく13本、歯骨におそらく13本の歯がある。上顎骨は背腹に高く、大きな癒合した歯間板が発達している。前眼窩窓の前方にはmaxillary fenestraなどの穴はない。ほとんどの仙前椎(頸椎と胴椎)が後凹型で、プレウロシールがあり、腹側に強いキールがある。腸骨は長く、丈が低い。恥骨の先端は少し膨らんだブーツをなす。前肢は短く、特に前腕が短い。
Galton and Jensen (1979)が最初に記載したときには頭骨を構成する骨は見つかっていなかったが、その後かなりの骨が発見され、頭骨の復元がなされた。Britt (1991)の時点で実際に発見されている骨は、前上顎骨、上顎骨、涙骨、後眼窩骨、頬骨、方形骨、歯骨の前半部分である。これらの骨はいずれも関節状態では見つかっていないが、涙骨以外は同一個体または同じ大きさの2個体のものと考えられている。特に前上顎骨と上顎骨はぴったりとフィットすることから、左右の頬骨は重なって見つかったことから、同一個体と考えられる。しかし上顎骨の後端は欠けていて涙骨や頬骨とつながっていないので、吻の長さや頭骨の長さと高さの比率は、ある程度誤差を含むはずである。展示された復元骨格の頭骨は、Britt (1991)の復元図にかなり忠実につくられている。そして、下顎の後方の骨(上角骨、角骨、関節骨など)は見つかっておらず、推定で作られている。下顎に不自然さを感じたのは妥当だったわけである。復元された頭骨の長さは1180 mm、高さは450 mmに達する。
前上顎骨はよく保存されており、体部は平行四辺形である。最近は図鑑等でも表現されているように、トルボサウルスの吻の先端(鼻孔より前方)は斜めになっている。前上顎骨の腹側縁と前縁のなす角度(premaxillary angle)は、トルボサウルスでは約55°であり、ケラトサウルスでは約80°、アロサウルスでは約72°であるという。この角度が小さいことはメガロサウルス類に共通した特徴のようで、エウストレプトスポンディルスやドゥブレウイロサウルスでもそうなっている。
多くの獣脚類では前上顎骨に4本の歯があるが、トルボサウルスの前上顎骨には3本しかない。これは他にはケラトサウルスにしかみられない特徴である。発見された前上顎骨には萌出した歯は保存されていないが、歯槽の形から前上顎骨歯はかなり大きく、断面は楕円形で、上顎骨や歯骨の歯とあまり変わらない形だったらしい。また前上顎骨歯は、ケラトサウルスと同様に互いに重なって並んでいる。
上顎骨は鼻骨突起の半分と後方の部分が欠けていたが、腹側縁はわずかにカーブしている。萌出した歯は保存されていないが、10個の歯槽が保存されており、部分的に欠けた11個目がある。全体の形から上顎骨歯は約13本と推定される。2番目から6番目の歯は大きく同じくらいのサイズで、他はより小さい。鼻骨突起の側面にある稜線が下方へ、次いで後方へカーブして前眼窩窩の前縁のカーブとなるが、トルボサウルスではこの下方へ曲がり始める位置(よってコーナーの位置)がアロサウルスやケラトサウルスよりも高いという。またアロサウルスでは前眼窩窩にmaxillary fenestraがあるが、トルボサウルスにはない。ただしトルボサウルスでは貫通した穴はないが、かなり深い窪みdepressionはある。この状態は獣脚類の原始的な状態をとどめているのだろうとしている。メガロサウルス類の中でもドゥブレウイロサウルスやアフロヴェナトルでは前眼窩窩に2つの穴がある。
涙骨の前方突起(鼻骨突起)は先細りで前端には指状の突起があるが、腹方突起(頬骨突起)の先端は幅広い扇形に拡がっている。背後方部分には、多くの獣脚類のものよりも小さい、粗面のある突起があるが、いわゆる涙骨角というほどの突起はない。また涙骨の頬骨突起は2枚の板を重ねたような構造をしているが、アロサウルスでは外側の板lateral bladeの方が幅広いのに対して、トルボサウルスでは内側の板medial bladeの方が幅広い。後眼窩骨の頬骨突起はアロサウルスよりも長く、上部2/3の断面はL字型で、下部1/3の断面はU字型である。
下顎の歯骨は、歯の生えた部分の大部分が保存されており、最初の9個の歯槽がよく保存され不完全な10個目がある。1本の萌出した歯といくつかの萌出していない歯がある。正確な歯の数は不明だがおそらく約13本と推定される。歯骨の上縁はかなりまっすぐで、下縁はゆるやかにカーブしている。歯骨の側面には、上縁に平行に一列の孔が並び、他にも歯骨の前端付近に多くの孔がある。1番目の歯槽には成長中の歯があり、その形態をみるとアロサウルスのように断面がD字型ではなく、またケラトサウルスのような内側の縦の稜線もない。
コメント ( 1 ) | Trackback ( 0 )
ドゥブレウイロサウルス1
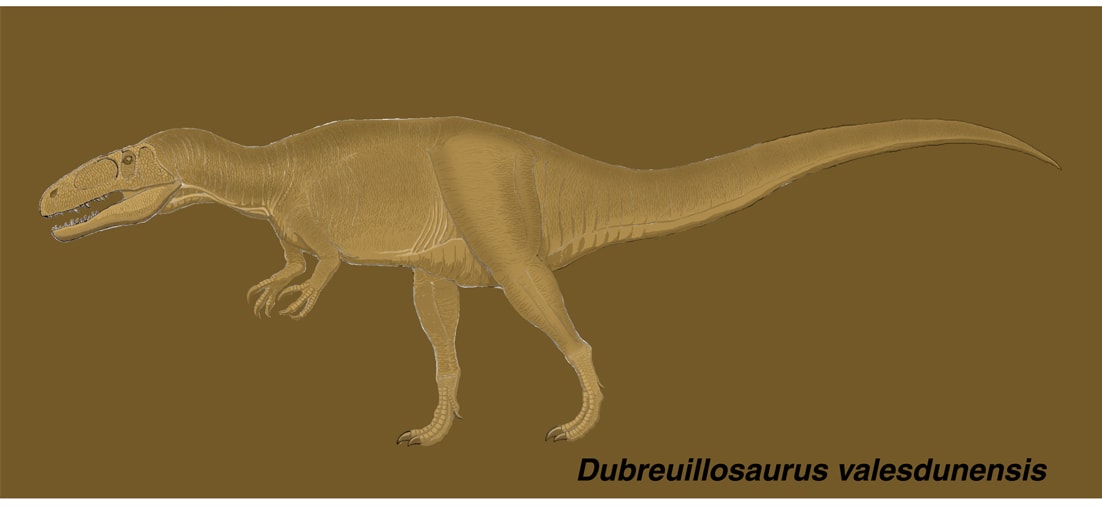
ドゥブレウイロサウルスは、ジュラ紀中期バス期にフランスのノルマンディー地方に生息したメガロサウルス類である。1994年にカルケール・ドゥ・カイン層から発見され,2002年に疑わしいポエキロプレウロン属の新種として報告されたが,その後尾椎の特徴などがポエキロプレウロンとは異なることがわかり、2005年に新属として記載された。頭骨の大部分と体の骨格の一部が発見されている。頭骨の研究から、メガロサウルス科の中でもエウストレプトスポンディルスやアフロヴェナトルと最も近縁とされている。
大型の獣脚類としては、かなり頭骨が長く丈が低い。復元された頭骨の長さ50cmに対し、眼窩の位置での高さは15cmである。このように頭骨の長さが高さの3倍以上あるのは、他にはスピノサウルス科とアフロヴェナトルくらいであるという。頭頂骨にnuchal crestがないため、側面からみると頭頂骨が見えない。上顎骨にはpromaxillary fenestra とmaxillary fenestra がある。上顎骨の鼻骨突起(後背方突起)の前縁のラインが途中で屈曲している。この屈曲は獣脚類の中で最も顕著であるという。
涙骨,前頭骨,鼻骨のいずれにも角やとさかなどの装飾はなく、シンプルな頭部である。(発見された涙骨は一部欠けていたが、涙骨角はないという。)トルボサウルス、アフロヴェナトル、エウストレプトスポンディルスと同様に、涙骨の鼻骨突起の前端は先細りになっているのに対し、頬骨突起(腹方突起)の先端は扇形に広がっている。下眼窩突起はない。進歩した獣脚類と異なり、後眼窩骨の腹方突起が眼窩の腹側縁まで伸びている。またトルボサウルス、アフロヴェナトル、エウストレプトスポンディルスと同様に、後眼窩骨の腹方突起は横幅が広く断面がU字型である。
頭骨以外の骨格は断片的で、正確なプロポーションは不明である。前肢は1個の末節骨しか見つかっていない。その形状はバリオニクスに似ているという。後肢は大腿骨、腓骨の一部と1個の中足骨、1個の趾骨がある。スピノサウルス上科としてはユニークな特徴として,大腿骨の軸に対して大腿骨頭が90度以上に曲がっている(はさまれる角度が90度以下)。これは、ケラトサウルスやディロフォサウルスのような、より原始的な獣脚類と同様であるという。
この恐竜はバリオニクスほどではないが,確かに顔が長く,もう少しでワニ顔になりそうな感じもある。実際にメガロサウルス類はスピノサウルス類と近縁である(スピノサウルス上科)。またポエキロプレウロンもドゥブレウイロサウルスも、マングローブの茂るような浅い海に面した海岸地帯に棲んでいたと考えられている。さらにポエキロプレウロンの腹部からは、胃石と思われる10個の小石とともに、当時のサメの一種ポリアクロドゥスの歯が見つかっていることから、メガロサウルス類は少なくとも食料の一部として,魚を食べたと考えられている。メガロサウルス類は海辺を徘徊しながら、陸上動物、翼竜、魚類,それらの死体など、利用できるものは何でも食べる肉食動物であったと想像される。これらとの共通祖先の中から,より魚食に適応して特殊化したスピノサウルス類が進化してきたというのは、ありそうなシナリオである。
参考文献
Allain, R. 2002. Discovery of megalosaur (Dinosauria, Theropoda) in the Middle Bathonian of Normandy (France) and its implications for the phylogeny of basal Tetanurae. Journal of Vertebrate Paleontology 22:548-563.
Allain, R. 2005. The postcranial anatomy of the megalosaur Dubreuillosaurus valesdunensis (Dinosauria, Theropoda) from the Middle Jurassic of Normandy, France. Journal of Vertebrate Paleontology 25:850-858.
コメント ( 0 ) | Trackback ( 0 )
| 次ページ » |