獣脚類を中心とした恐竜イラストサイト
肉食の系譜
カレンダー
| 2025年3月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | ||||||||
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| 9 | 10 | 11 | 12 | 13 | 14 | 15 | ||
| 16 | 17 | 18 | 19 | 20 | 21 | 22 | ||
| 23 | 24 | 25 | 26 | 27 | 28 | 29 | ||
| 30 | 31 | |||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
スピノサウルス類の骨密度

スピノサウルスとスコミムスでは骨の組織像が全然違うということは2014年にわかっていたわけだから、今回新しいのはバリオニクスのデータである。スピノサウルス類の中でもほとんど同じ体形のバリオニクスとスコミムスで生態が異なる、というのが面白いところである。
陸生脊椎動物である羊膜類が二次的に水中生活に戻る現象は、独立に30回も起きており、非常によくみられることである。その中で非鳥型恐竜は例外的で、ほとんど陸上生活に限られるとされてきた。その概念を打ち破ったのが、スピノサウルスのネオタイプの発見である。
水中での摂食subaqueous foragingや深い潜水deep divingのような水中生活への適応は、ボディプランの基本的な変更を伴う大きな進化的移行である。にもかかわらず、クジラ類や海生爬虫類のように高度の水生適応を示すグループでさえ、この変形には数百万年かかっている。いくつかの現生種や、最も特殊化した水生動物の系統でも初期の化石種では、比較的わずかな骨格上の変化しか示さない。例えばカバやごく初期のクジラ類などである。よって、現在陸生だったと考えられている恐竜の中にも、実は水生適応の初期段階にあったり、半水生だったりした種類がいるかもしれない。
骨密度の増加は、水中生活への適応として現生の羊膜類に広く生じており、ワニ形類、鳥類、海生爬虫類、クジラ類のような絶滅した四肢動物で水中生活を示唆するのに用いられてきた。骨密度は従来、1つの系統の中で古生態を考察するのに用いられてきたが、骨密度が水生適応の指標として有用であるかどうか評価するためには、もっと系統的に広範な検討が必要である。
そこでFabbri et al.(2022) は、206の現生および174の絶滅した羊膜類について、大腿骨の骨幹と肋骨の近位部の骨密度を定量化した。これらのデータは(1)水中摂食するかどうか(2)飛行するかどうかなどのスコアをつけられた。鳥類では飛行もするし潜水もする種もあれば、飛行できず潜水する種もある。データの中には議論の余地なく水生である現生種と絶滅種、例えば海生哺乳類(クジラ類、鰭脚類)、主竜類以外の海生爬虫類(魚竜類、鰭竜類、モササウルス類)、水生の主竜類(メトリオリンクス類、現生ワニ類)、潜水する鳥類(ペンギン、アビ、カイツブリなど)も含まれている。
最も相関が高かったのは骨密度と水中摂食で、つまり羊膜類全体で“頻繁な水中摂食”は、大腿骨と肋骨の骨密度の増加と相関していた。“頻繁でない水中摂食”や水面上からの摂食は、骨密度の増加と相関していなかった。これは渉禽類(サギ、ペリカン、フラミンゴなど)は陸生の動物と同様の骨密度をもつことと一致する。
ゾウや竜脚類のような巨大な陸上動物では、大腿骨の骨密度が増加する傾向があった。また魚竜類、モササウルス類、クジラ類、アザラシのように深く潜水する動物では、浅く潜水する水中摂食者と比べて低い骨密度を示した。これらの深く潜水する動物では緻密骨が海綿骨に置き換わり、多数の骨梁や血管で占められている。これは水圧への適応や代謝の増加と関連して説明されている。よって高い骨密度は、水生適応の初期段階の良い指標となるが、水面を歩く動物、深く潜水する動物、陸生動物を区別することはできないと考えられた。
著者らの解析の結果、非鳥型恐竜の中でスピノサウルス類だけが、はっきりと水中摂食者と判定された。スピノサウルス類の中でも多様性があり、スピノサウルスとバリオニクスは水中摂食者と予測されたが、スコミムスの骨密度は他の陸生の獣脚類と同様で、水中摂食者でないと判定された。スコミムスの大腿骨はティラノサウルス、ティラノティタン、トルボサウルスなどの大型獣脚類と似ていたが、スピノサウルスとバリオニクスは大きく異なっていた。
オルニトミムス類、ハルシュカラプトル、鳥脚類など他の非鳥型恐竜はすべて、中空の骨髄腔をもち、水中摂食者とは予測されなかった。それに対してバリオニクス(背側肋骨、肩甲骨、恥骨、座骨、大腿骨、腓骨)とスピノサウルスのネオタイプ(背側肋骨、胴椎と尾椎の神経棘、大腿骨、脛骨、腓骨、手の指骨)では多数の骨に高い骨密度が観察され、水中摂食者という予測は確実と思われた。
系統的な比較から、スピノサウルス類の祖先は水中摂食をしていたと考えられ、スコミムスは二次的に高い骨密度を失ったと考えられる。骨密度が高くないからといって、スコミムスが水辺の環境に依存していたことを否定するものではない。長い吻や円錐形の歯などの形態学的特徴は主に魚食性であったことを示している。従来から想定されたように、スコミムスは河岸あるいは浅瀬に立って魚を捕らえるハンターだった可能性が高い。形態学的には同じように見えるバリオニクスとスコミムスで、潜水するものとしないものという大きな生態学的違いがあったことになるが、これはバリオニクス亜科に限ったことではなく、ウ科やカバ上科にもみられるという。これは河川や沼などの水辺がまばらに分布していたなどの環境要因が関係しているかもしれないとある。
つまりバリオニクス「重い爪」は爪だけでなく、体の骨も重かったというオチである。
参考文献
Fabbri, M., Navalón, G., Benson, R.B.J. et al. Subaqueous foraging among carnivorous dinosaurs. Nature 603, 852–857 (2022). https://doi.org/10.1038/s41586-022-04528-0
コメント ( 0 ) | Trackback ( 0 )
ケラトスコプス・インフェロディオス

後眼窩骨を赤くしたいがために、全身描いたようなものだ。バリオニクス30個体とスコミムス40個体で種内変異を研究したら、どうなるだろう。
コメント ( 0 ) | Trackback ( 0 )
ケラトスコプスとリパロヴェナトル

Copyright 2021 Barker et al.
本当に別属とする必要があるのだろうか。例えばスコミムスを含めてバリオニクス属の新種ではいけないのだろうか。種内変異の幅を知らないうちに形態を細かく解析すると、個体ごとに新属新種になったりしないのだろうか。そういう感想である。今回の2つの新標本はバリオニクスよりもスコミムスに近い点があり、バリオニクスとスコミムスを属として残すのであれば、新属になるということか。
ケラトスコプスとリパロヴェナトルは、白亜紀前期バレミアンに英国南部のワイト島に生息したバリオニクス亜科のスピノサウルス類で、2021年に記載された。
英国では、恐竜化石を産する前期白亜紀のウェルデン層群がイングランド南東部とワイト島に露出している。バリオニクス・ワルケリのホロタイプ標本は、イングランド南東部サリーのUpper Weald Clay Formation(バレミアン)から発見された。この発見によりウェルデン層群の様々な分離したスピノサウルス類化石が、バリオニクスと呼ばれていた。スピノサウルス類の化石はワイト島のWessex Formation(バレミアン)からも知られていたが、これまで報告されたのは分離した歯と1個の胴椎であった。これらはバリオニクスまたはその近縁種と考えられ、cf. BaryonyxやBaryonyx sp.などと称された。これらの分離した歯とバリオニクスのホロタイプの歯では、エナメルの装飾に異なる特徴がみられることから、別のスピノサウルス類の可能性も考えられたがはっきりしなかった。
今回、新たに2つのスピノサウルス類の断片的な化石標本が、ワイト島南西部のChilton Chineから発見された。この2つの標本は、驚くべきことにほぼ同時代のバリオニクス・ワルケリとは異なり、また互いに異なっていたので、ケラトスコプス・インフェロディオスCeratosuchops inferodiosとリパロヴェナトル・ミルネラエRiparovenator milneraeと命名された。ほぼ同時代の同一地域に、複数のスピノサウルス類が共存していたことになるという。
ケラトスコプス・インフェロディオスCeratosuchops inferodiosの属名は「角のあるワニの顔」で、種小名は「地獄のサギ」である。この場合の角というのはバリオニクスのような正中のトサカではなくて、後眼窩骨の突起やごつごつした粗面をさす。
ケラトスコプスのホロタイプ標本は、左右の前上顎骨、後方の前上顎骨の断片、ほとんど完全な分離した脳函からなる。また参照標本として右の後眼窩骨がある。
他のバリオニクス亜科と異なるケラトスコプスの特徴は、前上顎骨の外鼻孔の前方に結節があること、基底後頭骨のsubcondylar recessの幅が狭く腹方に限られること、などの非常に細かい形質からなる。
リパロヴェナトル・ミルネラエRiparovenator milneraeの属名は「河岸のハンター」で、種小名はバリオニクスを研究したアンジェラ・ミルナー博士への献名である。
リパロヴェナトルのホロタイプ標本は、左右の前上顎骨、分離した脳函、左の眼窩前方の断片(部分的な涙骨と前前頭骨)からなる。参照標本として後方の鼻骨の断片と、連続した尾椎の一部がある。
他のバリオニクス亜科と異なるリパロヴェナトルの特徴は、眼窩の上の前前頭骨と前頭骨の後眼窩骨突起との間に切れ込みnotchがあること、顔面神経孔が深く陥没していること、subcondylar recessが深いこと、などの細かい形質からなる。
系統解析の結果、スピノサウルス類の単系は強く支持され、またスピノサウルス亜科とバリオニクス亜科の2分岐が支持された。バリオニクス亜科の中では、ケラトスコプスとリパロヴェナトルが姉妹群となり、これらとスコミムスを含めた新しいクレード、ケラトスコプス族Ceratosuchopsiniが認識された。ケラトスコプス族の特徴は、前頭骨の後眼窩骨との関節面が背腹に厚く、深い長軸方向の溝があること、上側頭窩の前縁が明確で強くカーブしていること、などであるという。
分類学的な解釈については、これらのスピノサウルス類の頭骨の細かい形質の違いは、個体変異や成長段階による差異に関するものを含んでいることは認めている。これを正しく評価するためにはもっと多数のサンプルが必要であり、それが不可能である現在のところは、新しく2種を命名するのが妥当であるということをいっている。
今回の新種で注目される形質として、後眼窩骨の粗面がある。ケラトスコプスでは後眼窩骨の粗面の最後に、大きな突起bossがある。またリパロヴェナトルとスコミムスでも、前頭骨の後眼窩骨との関節面の形状が似ていることから、同様の後眼窩骨があったことが暗示される。このような頭部の装飾は、ディスプレイや種内闘争に役立った可能性がある。また今回の論文では個々の骨の詳細は記載していないが、リパロヴェナトルの尾椎では神経棘が比較的長く、丈の高い尾椎の起源がさかのぼることを示唆する。これまでは丈の高い尾椎はアプティアンのイクチオヴェナトルとスコミムスからとされていた。
英国のウェルデン層群のスピノサウルス類の歯化石には、2つ以上の形態型morphotypeがみられることから、複数のバリオニクス亜科の種が存在した可能性は以前から指摘されていた。ただしこのことは、複数の種がいたというよりも、スピノサウルス類の歯の異歯性の程度が著しいことを表すとも議論されていた。しかし前期白亜紀のイベリア半島からは複数のスピノサウルス類と多数の歯の形態型が知られており、今回のケラトスコプスとリパロヴェナトルの発見を考え合わせると、より複雑な状況が想像される。バレミアンのイベリア半島にはバリオニクス、ヴァリボナヴェナトリクス、カマリラサウルスが生息し、バリオニクス亜科とスピノサウルス亜科の両方を含むとされる複数の歯が見つかっている。ただしこれらの種類は今のところ異なる地層から知られており、実際にどの程度共存したのかは明確ではない。
ケラトスコプスとリパロヴェナトルの歯には、唇側と舌側にエナメルの装飾が観察される。これはこの地層から見つかっていた分離した歯と同じ特徴であり、バリオニクス・ワルケリの舌側に縦溝のある形態とは異なっている。今回の発見により、ワイト島のWessex Formationにバリオニクスそのものが存在したかどうかは、疑わしくなってきた。実際、過去にバリオニクスと呼ばれた分離した胴椎は、神経棘がより高いなどの異なる特徴をもっている。他の標本についても未同定のバリオニクス亜科あるいはスピノサウルス科と呼ぶのが良いだろうとしている。
古生物地理学的には、スピノサウルス亜科がゴンドワナ地域に分布し、バリオニクス亜科が主にローラシアに分布していることから、スピノサウルス類は最初はパンゲア全体に分布しており、大陸の分裂にともなってゴンドワナでスピノサウルス亜科が、ローラシアでバリオニクス亜科が分化した(スコミムスだけがアフリカに渡った)という考えもあった。
しかし今回の結果はスピノサウルス類のヨーロッパ起源を支持し、その後少なくとも2回、アフリカに渡ったことを示唆するとしている。1つはスコミムスであり、もう一つはスピノサウルス亜科の祖先の段階である。スペインのカマリラサウルスを基盤的なスピノサウルス亜科と考えると、状況を単純化することができるといっている。
参考文献
Barker, C.T., Hone, D.W.E., Naish, D. et al. New spinosaurids from the Wessex Formation (Early Cretaceous, UK) and the European origins of Spinosauridae. Sci Rep 11, 19340 (2021). https://doi.org/10.1038/s41598-021-97870-8
コメント ( 0 ) | Trackback ( 0 )
スピノサウルスについての議論
スピノサウルスについての議論は本当に尽きなくて、当分の間楽しめますね。
ナショジオの記事ではイブラヒム側とホルツ側の双方に取材しているが、確かにイブラヒム側のいうように、ホルツ側は水中捕食者の定義を狭くしすぎているというのはわかる。
私は東京港野鳥公園で、たまたまウが潜水して水面下で魚を追いかけるのを見たが、後肢で強力に推進しながら、水中で頭を魚に向かって突き出していた。長いS字状の首で。。。それで害鳥にされるほど繁栄している。アビやカイツブリも水中で捕食するのに何も困っていないようだし。
獣脚類のS字状の首は、主竜類以来の進化史の遺産(レガシー)だから、そう簡単に変更しにくいというのもあるだろう。そもそもスピノサウルスの祖先の原始的なスピノサウルス類の段階では、水辺で水面上から吻を突っ込んで捕食していたに違いないわけで、その時点でS字状の首は大変重宝していた形質のはずである。それを捨て去るにはよほどのメリットがないといけないのではないか。スピノサウルス類が獲得した長めのS字状の首を保持しているからといって、常に水面上から狩りをしたとも限らないのではないか。そういえば原クジラでも「サギのように首が長い」種類がいたような気がする。
スピノサウルスの尾が扁平でも、ワニほどの推進力はないともいっている。
原始爬虫類の基本形から進化したなら、ワニやモササウルスのように全身をくねらせるのが最適かもしれないが(ワニ類は地上性の2足・4足を経ているとしても)、獣脚類は胴を固定して2足歩行するのを基本として進化してきた。その条件下での水生適応の話だから、例えば後肢と尾を併用して泳いだのではないか。
生物の形態が、常に最適化された状態を表しているという概念も、100%ではないかもしれない。キノボリカンガルーは木登りが苦手なことで有名である。まだ木登りに十分適応していない状態ともいわれている。骨格の化石だけを機能形態学的に解析したら(そういう研究がされているかどうか知らないが)、この動物は木登りに最適化されていない。よって木には登らないはずだ、と結論されることはないだろうか。
ナショジオの記事ではイブラヒム側とホルツ側の双方に取材しているが、確かにイブラヒム側のいうように、ホルツ側は水中捕食者の定義を狭くしすぎているというのはわかる。
私は東京港野鳥公園で、たまたまウが潜水して水面下で魚を追いかけるのを見たが、後肢で強力に推進しながら、水中で頭を魚に向かって突き出していた。長いS字状の首で。。。それで害鳥にされるほど繁栄している。アビやカイツブリも水中で捕食するのに何も困っていないようだし。
獣脚類のS字状の首は、主竜類以来の進化史の遺産(レガシー)だから、そう簡単に変更しにくいというのもあるだろう。そもそもスピノサウルスの祖先の原始的なスピノサウルス類の段階では、水辺で水面上から吻を突っ込んで捕食していたに違いないわけで、その時点でS字状の首は大変重宝していた形質のはずである。それを捨て去るにはよほどのメリットがないといけないのではないか。スピノサウルス類が獲得した長めのS字状の首を保持しているからといって、常に水面上から狩りをしたとも限らないのではないか。そういえば原クジラでも「サギのように首が長い」種類がいたような気がする。
スピノサウルスの尾が扁平でも、ワニほどの推進力はないともいっている。
原始爬虫類の基本形から進化したなら、ワニやモササウルスのように全身をくねらせるのが最適かもしれないが(ワニ類は地上性の2足・4足を経ているとしても)、獣脚類は胴を固定して2足歩行するのを基本として進化してきた。その条件下での水生適応の話だから、例えば後肢と尾を併用して泳いだのではないか。
生物の形態が、常に最適化された状態を表しているという概念も、100%ではないかもしれない。キノボリカンガルーは木登りが苦手なことで有名である。まだ木登りに十分適応していない状態ともいわれている。骨格の化石だけを機能形態学的に解析したら(そういう研究がされているかどうか知らないが)、この動物は木登りに最適化されていない。よって木には登らないはずだ、と結論されることはないだろうか。
コメント ( 0 ) | Trackback ( 0 )
スピノサウルスは沈まない、浮いても不安定、重心は腰に近い (3)

Copyright 2018 Henderson
浮いてもすぐ横倒しに
Henderson (2018)は、アリゲーターとスピノサウルスについて、それぞれ胴の重心近くの断面モデルを作成した。アリゲーターの胴の断面モデルは円に近い形に、スピノサウルスの胴の断面モデルは縦に長い楕円形になった。
著者はこれらのモデルについて、重心(CM, center of mass)、浮心(浮力中心)(CB, center of buoyancy)、メタセンタ高さ(MC, metacentric height)を計算した。メタセンタ高さとは造船学、船舶工学の用語で難しいが、安定性に関する概念のようだ。水中に浮かんでいる物体について、メタセンタ高さMCが下にあり、重心CMが上にあるのは不安定な状態であるという。
アリゲーターの胴の断面モデルでは、CM 、CB 、MCの3つがほとんど一致しており、これは安定な状態である。一方でスピノサウルスの胴の断面モデルでは、CMがMCよりも22 cm上方にあり、明らかに不安定な状態であるという。

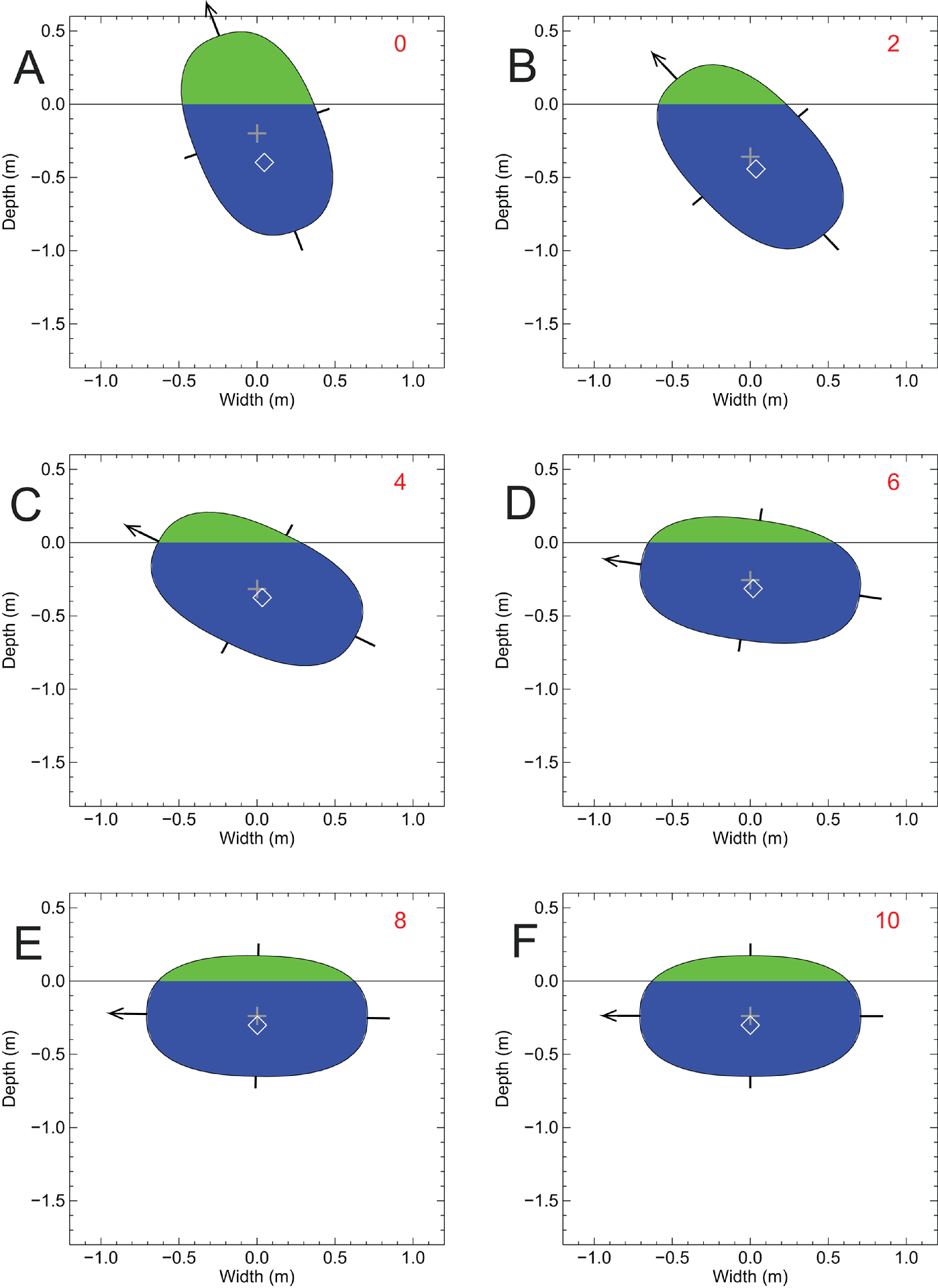
さらに側面方向の安定性をテストするため、Henderson (2018)はそれぞれの断面モデルを20°傾けて、重力と浮力が平衡に達するまで動き続けるシミュレーションを行った。するとアリゲーターの胴の断面モデルは、徐々に傾きが回復し、最終的には元の上端(背中)が上にくる位置で安定に達した。ところが、スピノサウルスの胴の断面モデルは20°傾けられるとそのまま横に倒れてしまい、回復することなくその状態で平衡に達してしまった。このあたりは直観的には誰でも思うことであるが、実際にモデル化して重力と浮力を計算し、結果を視覚的に見せるところが重要なのだろう。工学系の人にはそれほど難しくはないのだろうが、このくらいやらないと論文のデータとして示せないということだろう。
このことは、水に浮かんだスピノサウルスの胴体はちょっと力が加わると傾きやすく、姿勢を保つために絶えず四肢を動かしていなければならないことを示唆するといっている。これでは半水生生活に適しているとはいえないというわけである。
新復元について
Henderson (2018)は容赦なくIbrahim et al. (2014)の新復元にも突っ込み続ける。スピノサウルスの新復元は、複数個体の骨格をスケールを合わせて合成したものであり、特に後肢の骨格は、脊椎全体と同一個体のものではない。復元の過程で誤りがあれば、胴体と後肢の比率などに大きく影響するだろうという。後肢の重量の体重に対する比率をグラフにプロットし、スピノサウルスの新復元の後肢の小ささは、他の5種の獣脚類とはかけ離れていることを示している。
また現生の半水生のワニは、祖先の陸上性のワニ類に比べて短い四肢を持っているが、この場合後肢だけでなく、前肢も短くなっている。これは水中を泳ぐ際の抵抗を減らすとともに、推進力として中軸の胴体の筋肉系の優位性を示している。スピノサウルスの新復元の後肢の縮小が、水中生活への適応を表すとすれば、ワニ類のように前肢も小さくなってよさそうなものである。しかし新復元では前肢は地上に届くほどの長さとなっている。前肢の骨は由来がバラバラであり、一部の骨は新しい標本と共に見つかっているが、他の骨のスケールをどのように合わせたのか明確でないといっている。つまり、暗に不自然ではないか、四足歩行のイメージありきで合わせたのではないかといっているのである。
Henderson (2018)は、スピノサウルスが大部分の時間を水中で過ごすほど水中生活に適応していたことを疑問視しているが、バリオニクスなどと同様に水辺に生息し、浅瀬に入るなどして魚を捕食していたことは否定していない。Ibrahim et al. (2014) の半水生説ではなく、他のスピノサウルス類と同じような水辺の生活をしていたという、もっと保守的な考えが依然として有力な可能性の一つであると締めくくっている。
ちなみに、スピノサウルスは水に浮くが、他の獣脚類と比べて特別なことは何もない、というくだりで軽いデジャヴを覚えた。吻部の神経血管網もそうであった。
基本的にコンピューター上の3次元モデルでの検証であるし、多くの仮定を重ねているので、本当にその設定で妥当なのか不安になるところはある。鳥類と同様の気嚢系があると設定して計算すれば、それは密度は小さくなるだろうし沈まないと出るだろう、という意見もあるだろう。
しかしIbrahim et al. (2014) の半水生説の問題点についていろいろと突っ込んでいるので、恐竜ファンの興味を引く大変面白い研究である。あのダックスフント型四足歩行を否定するまでには至らないとしても、二足歩行の復活を待ち望む一部の(少なからぬ)恐竜ファンにとって、希望の光は失われていないかもしれない。
了
コメント ( 0 ) | Trackback ( 0 )
スピノサウルスは沈まない、浮いても不安定、重心は腰に近い (2)
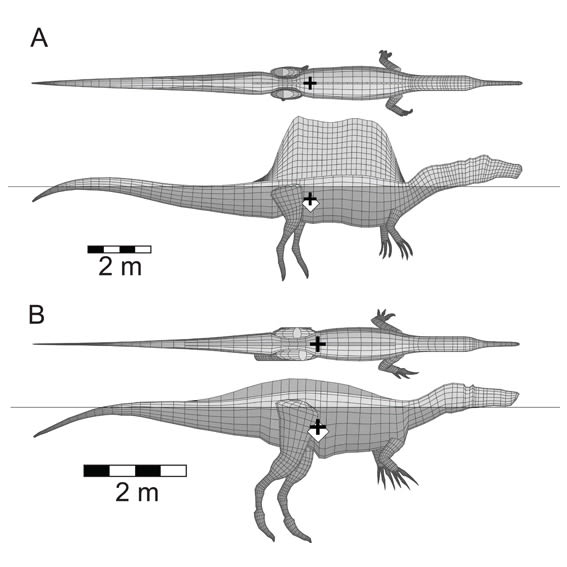
Copyright 2018 Henderson スピノサウルス類の3次元モデルを水に浮かべたところ
沈まないこと
Hendersonの過去の研究によると、アリゲーターは肺の空気の40-50%を排出すれば、沈むことができるという。一方、気嚢系をもつ獣脚類は沈めない可能性がある。スピノサウルスのモデルの体密度は833 g/lで、肺の空気を75%排出したとしても885 g/lとなる。これは水の密度1000 g/lよりも小さいので、依然としてスピノサウルスは浮いてしまうことになる。
現生の半水生の鳥類や爬虫類、たとえばペンギン、アビ、ウ、ウミヘビ、ウミイグアナ、ワニなどは皆、水中に潜って獲物の追跡などを行う能力をもっている。カワウソやカワネズミなどの半水生哺乳類も同様である。これらの動物にとって、完全に水中に潜れないことは大きな障害となると考えられる。スピノサウルスが水中に潜れないとすれば、水生の獲物を捕食する能力が大きく制限されることになり、スピノサウルスが半水生生活によく適応していたというIbrahim et al. (2014) の考えと矛盾する。
これらの獣脚類で、気嚢系の発達の程度が現生鳥類と同じと考えるのが過大なのかもしれない。Henderson (2018) は気嚢系がなかった場合についても検討している。この場合、中軸の胴体は均一で1000 g/lと設定し、四肢と帆はそのままで、肺もそのままとした。このモデルでは体密度は918 g/lとなり、肺の空気を75%排出すると986 g/lとなった。これでも水の密度1000 g/lよりも小さいので沈むことはできないと述べている。
Henderson (2018)の論調はこのように、密度が水よりも小さい=沈めない=潜れないといっているようにみえるが、このあたりには少し疑問がある。
ペンギンには他の鳥類にあるような気嚢系がなく、骨の密度が高くなっているという。コウテイペンギンのモデルの体密度は968 g/lで、肺の空気を90%排出すると989 g/lとなり水(ここでは海水)よりは小さい。しかし実際にはペンギンは、潜る前に息を吸い込むという。ペンギンはヒレ状の翼と強力な胸筋で、浮力を克服して水中に潜ることができるとある。ここで列挙された半水生の鳥類や爬虫類が、それぞれどうやって潜水しているのか、説明がない。実際に肺の空気を吐き出すことで比重を変えているのだろうか。特にもともと気嚢系をもっている鳥類が、どのように潜水しているのかについて、もっと説明がほしいところである。鳥類の生理学として研究されていそうな気がする。つまり、体密度が水の密度1000 g/lよりも小さいイコール潜水できない、ではないような気がする。
全体の論調としては、ワニの952 g/lやペンギンの968 g/lはぎりぎり浮いているレベルで、姿勢や運動によって容易に調節できるが、スピノサウルスの833 g/lは軽すぎて、潜ることが困難だろうということだろう。それは理解できる。半水生説の立場に立てば、スコミムスまでは気嚢系があったがスピノサウルスでは急激に退化していたという証拠があれば、なんとか潜れたかもしれない。
獣脚類の気嚢系が、鳥類と同じ程度で15%引きというのがどうかと思ったが、重心を計算するとちょうどいい位置にくるわけだから、それほど過剰な見積もりではないということだろう。二足歩行するために前半身をそのくらい軽量化しているわけである。そうすると、比重は小さくならざるを得ない。スピノサウルスについても、特別な根拠がない限り、他の獣脚類と同様と考えるのが基本ということだろう。
つづくかな
コメント ( 1 ) | Trackback ( 0 )
スピノサウルスは沈まない、浮いても不安定、重心は腰に近い (1)
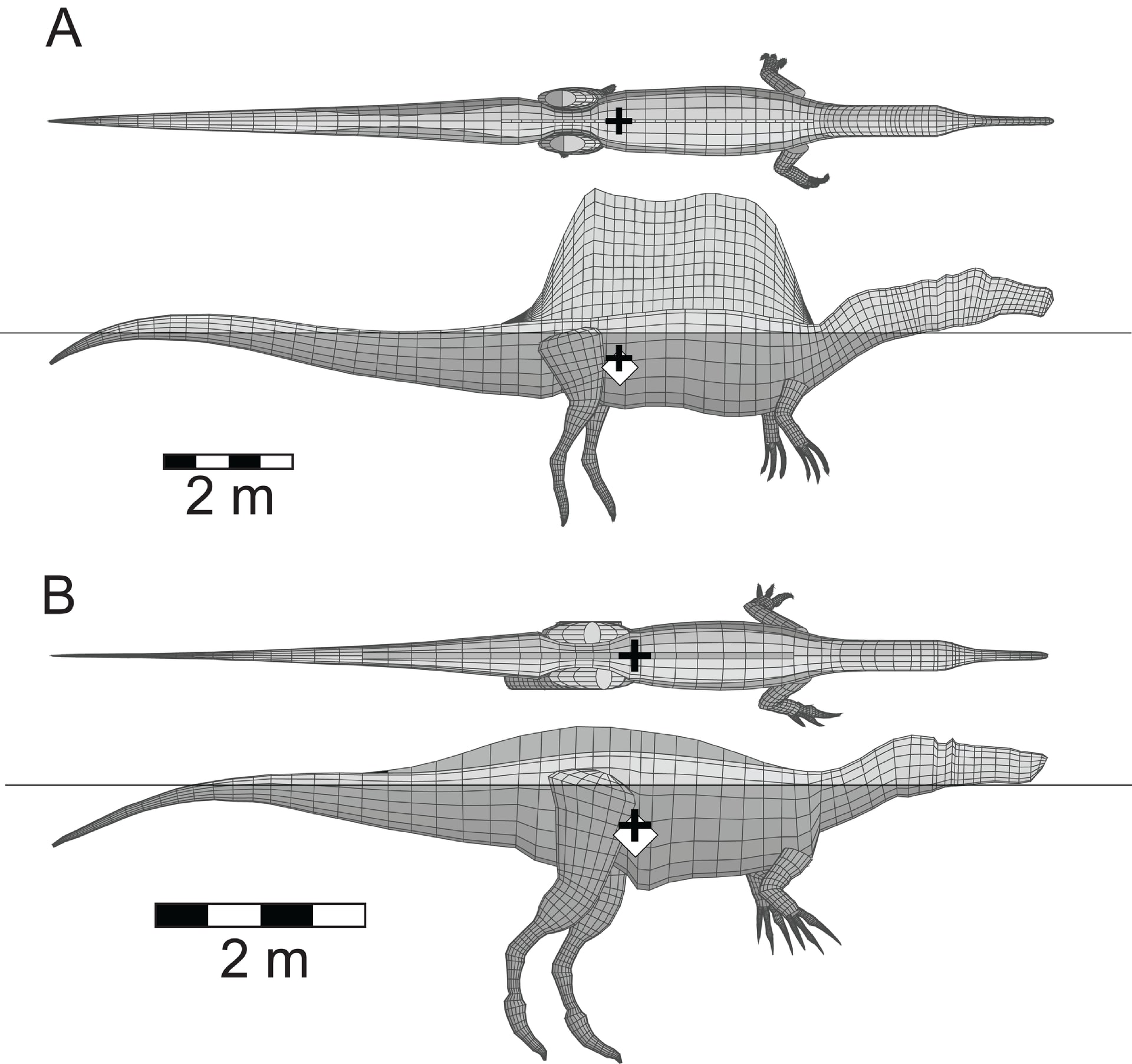
Copyright 2018 Henderson

大きい画像
謎の恐竜スピノサウルスの研究は、まだまだ話題が尽きないという感じである。
Ibrahim et al. (2014) の半水生適応説、つまりスピノサウルスは四足歩行で大部分の時間を水中で過ごす半水生生活を送っていたという考えに対して、ロイヤルティレル古生物学博物館のHenderson (2018) は大きな疑問を呈する論文を報告した。これは、半水生適応説について誰もが疑問に思う点を、モデルを使って定量的に表現したような研究である。あの帆ではどうみても水中を泳ぐのに邪魔ではないのか、とは恐竜ファンの誰もが思っただろう。
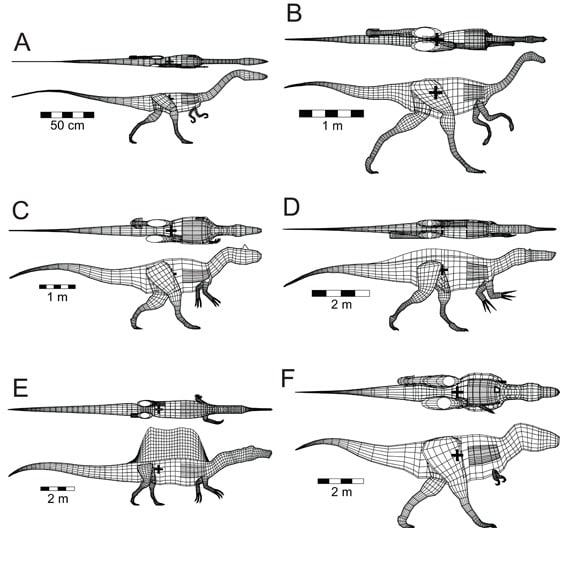
Henderson (2018)はいくつかの問題点について検証するため、体の部位ごとの密度の違いや肺、気嚢系を考慮したスピノサウルスの3次元デジタルモデルを作成し、このモデルの浮遊に関する性質を、専用のソフトウェアを用いて解析した。比較のため、近縁のスコミムス(ここではバリオニクス)や他の獣脚類ティラノサウルス、アロサウルス、ストゥルティオミムス、コエロフィシスの3次元デジタルモデルも同様に作成した。さらにモデル化の妥当性を検証するため、アリゲーターとコウテイペンギンのモデルも作成し、比較している。
スピノサウルスの3次元モデルは、Ibrahim et al. (2014)のFig. S3の新復元図からそのままとってコンピューター上で立体化している。他の獣脚類はグレゴリー・ポールなどいくつかの復元図をもとに、肉付けして3次元化している。
Henderson (2018)は浮力や安定性などを計算するために、これらの3次元モデルの密度分布を考慮している。四肢を除いた中軸の胴体の組織密度は、基本的に水と同じ1,000 g/l と設定されたが、獣脚類の解剖学的特徴を反映して調整された。現生鳥類の体内の気嚢系は、中軸の胴体体積の約15%を占めることから、これを獣脚類のモデルに応用している。絶滅した獣脚類の骨格にも広範な含気性の証拠があり、鳥類と同じような気嚢系をもっていたと考えられることから、腰から前方の胴体の密度は15%引いて、850 g/lとした。つまり腰部、胴部、頸部の気嚢の分である。気嚢の相対的な割合はどの獣脚類も同じと想定している。頭骨にも含気性があり、鼻腔や口腔もあることから、頭部も同様に850 g/lとした。さらに、肺の分がある。現生爬虫類のデータに基づいて、どの獣脚類モデルも、肺の体積は中軸胴体体積の約9%と設定された。最後に四肢は、やや骨が多いということで1,050 gm/lと設定された。
スピノサウルスに特徴的な背中の帆については、特別な注意を払って細かく計算している。Ibrahim et al. (2014)の復元図から神経棘の輪郭をとり、側面の面積で骨と軟組織の比率を計算したり、皮膚の厚みを1 cmと仮定するなどいろいろ計算した結果、帆の総重量は335 kgと推定され、これは 中軸胴体重量の7.5%を占めると算出された。
重心の位置
6種の獣脚類の3次元モデルで重心の位置を計算したところ、どの獣脚類でも重心は腰関節の少し前方にきた。これは二足歩行するときの後肢の範囲内に収まっているので、安定して歩行できる位置である。これはスピノサウルスにも当てはまった。重心の位置はスピノサウルスでもスコミムスとほとんど変わらず、腰の少し前方になった。
つまりスピノサウルスの新復元では、重心は胴の中央付近にくるというIbrahim et al. (2014)の結果と異なり、今回の研究では重心はもっと腰関節に近い位置になった。このことからスピノサウルスは後肢が短いけれども、陸上を歩行できたことが示唆されるとHenderson (2018)はいっている。
ただ筆者の感想としては、前後方向の重心の位置としてはそうかもしれないが、新復元のきゃしゃな後肢では、とてもスピノサウルスの巨体を支えられるようには思えないが。では四足歩行かというと、前肢は(ほとんど見つかっていないともいえるが)歩行に用いるようにはできていないので、それも疑問な気がする。実は後肢は別個体で、本当はもっと大きく、太く頑丈な後肢であれば納得がいく。
また背中の帆は重心を前方にずらすように働くようにみえるが、あまり影響がないのが意外である。気嚢系や肺の減量効果が大きいということかと思ったが、実はHenderson (2018)はそれについて検討していて、気嚢系や肺がなくても重心は腰関節の前方にくるという結果を載せている。ただしこの計算には背中の帆は含まれていないのはなぜなのか、よくわからない。この図をみると、新復元の尾が長いことが効いているように思える。
次にHenderson (2018)は、6種の獣脚類の3次元モデルをコンピューター上で水に浮かべてみた。その結果すべての獣脚類は、少し頸を上向きに曲げるなどすれば、頭部とくに鼻孔を水面上に出したまま、浮くことができた。多くの陸上動物が水面に浮いて泳ぐことができることを考えると、このことは不思議ではない。スピノサウルスもスコミムスも頭を出して浮くことができるが、陸生の獣脚類もみな同様であって、スピノサウルスに特別な点は何もなかった。なお、背中の帆は水面上に出ていることがわかった。
著者は、同様に作成したアリゲーターとコウテイペンギンの3次元モデルも水に浮かべて、それらが実際に野外で観察される水面上の休息姿勢とよく一致することを確認している。つまり水面下に沈む部分の割合などである。
つづく
参考文献
Henderson (2018) A buoyancy, balance and stability challenge to the hypothesis of a semi-aquatic Spinosaurus Stromer, 1915 (Dinosauria: Theropoda). PeerJ 6:e5409; DOI 10.7717/peerj.5409
コメント ( 0 ) | Trackback ( 0 )
イリタトル
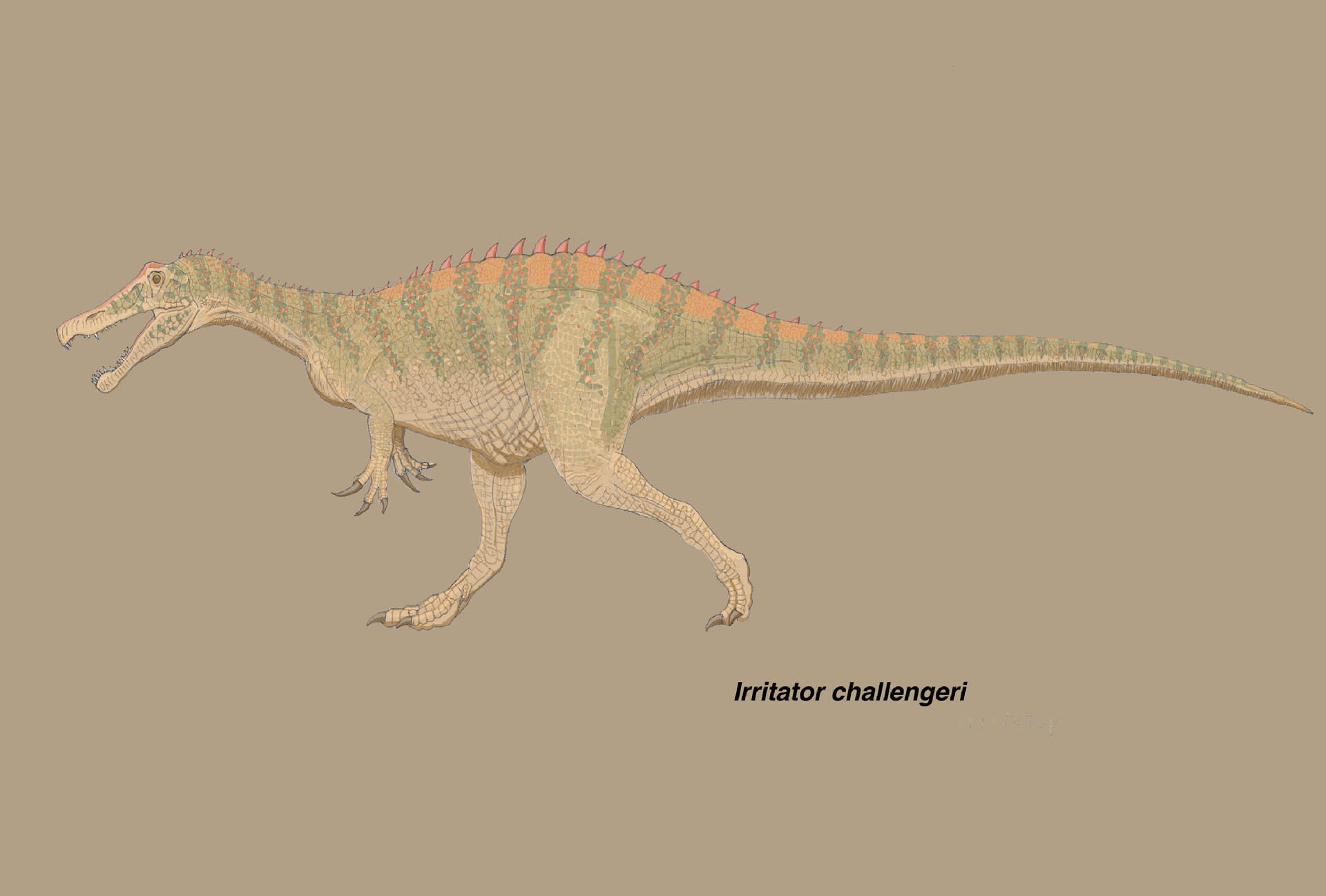
Sales and Schultz (2017) による改訂された特徴では、イリタトルはバリオニクス、スコミムス、クリスタトゥサウルスとは、歯に鋸歯がないことと上顎骨の歯槽の数が約半分しかないことで区別される。また“スピノサウルス” MSNM V4047や“スピノサウルス・マロッカヌス”MNHN SAM 124とは、外鼻孔がより大きく、より前方にあること、外鼻孔の前縁に前上顎骨が面していることで区別される。さらにイリタトルの固有形質としては、左右の鼻骨が結合してできた正中のとさかがあり、その後端が前頭骨の上で取っ手状knob-likeの突起をなすことである。
スピノサウルス類は、ディスプレイに利用できる構造を2つ持っている。頭部の鼻骨にある小さい“とさか”と、背中の長く伸びた“帆”である。これらについて、Hone and Holtz (2017) は次のように考察している。
頭部の小さいとさかは多くの獣脚類にみられるもので、鼻骨や涙骨など、眼窩の近くにある。捕食者である獣脚類が比較的小さなとさかを持っていることは、社会的・性的シグナルとしての役割と、獲物に警戒させない(察知されない)ための要請との取引の結果(trade-off 、妥協)と考えられる。あまり目立つ構造や色をしていると、獲物に察知されやすいわけである。そこでスピノサウルス類に背中の帆が発達した理由として、これらは水面上から水中に吻を突っ込んで魚などを捕食することをあげている。背中の帆は水面上にあって水中の魚には見えないというわけである。
なるほど、地上性の大型肉食恐竜が大きな帆をもっていたら、遠方からでも獲物に察知されてしまうかもしれない。しかしアクロカントサウルスやヤンチュアノサウルスなども長めの神経棘をもつので、基盤的なテタヌラ類に共通した別の理由があるのかもしれない。また、系統に関係なく、巨大化する際の骨の成長に関わる何らかの傾向のような気もする。
参考文献
Hone DWE, Holtz TR. (2017) A century of spinosaurs-A review and revision of the Spinosauridae with comments on their ecology. Acta Geol Sin-Engl. 91: 1120-1132.
コメント ( 0 ) | Trackback ( 0 )
ブラジルのスピノサウルス類の再検討とスピノサウルス類の頭蓋の進化(下)

Copyright 2017 Sales and Schultz
論文の復元頭骨図は、イリタトルと比べてアンガトゥラマの方が少し大きいことを表している。オクサライアは時代も後であり、アフリカのスピノサウルス亜科と近縁と考えているので、スピノサウルスの復元頭骨に当てはめるとこうなるのかもしれない。
円錐形の歯、細長い吻、後退した外鼻孔など、スピノサウルス類の多くの頭蓋と歯の特徴は半水生の生活と関連している。また歯や骨の同位体組成の分析や組織学的解析からも、スピノサウルス類は半水生や水生の動物と近いことがわかっている。しかし、同位体組成のデータは、スピノサウルス類の中でも、より陸生に近いものとより水生に近いものがいたことを示している。またスコミムスとスピノサウルスのネオタイプの肢の骨の組織切片からも、水中環境への適応の程度に差があったことが示されている。
このようなスピノサウルス類の中での生態の多様性が、外鼻孔の状態とも関連していると思われる。スピノサウルス類の中で、外鼻孔の位置と大きさはさまざまである。外鼻孔が小さいことは嗅覚の重要性が低いことを示し、外鼻孔が大きいことは嗅覚の重要性が高いことを示すと考えられる。外鼻孔が最も小さい“スピノサウルス”ではまた、外鼻孔が最も後方に移動している。外鼻孔が後方にあることは、嗅覚刺激の収集に向いていない。またもう一つの観点として、外鼻孔と嗅球の間のスペースには鼻腔があり、間接的に嗅上皮の表面積を反映している。種々のスピノサウルス類の頭骨を比較すると、外鼻孔の位置が前方にあるほど鼻腔が大きく、嗅上皮の表面積が大きいことになるわけである。
魚食性の捕食者とくに半水生の四肢動物にとって、嗅覚は主要な感覚ではない。これらの動物にとって、嗅覚以外の感覚が重要である。ワニ類は吻部に機械受容器をそなえており、これで水の動きを感知する。鳥類と翼竜類では主に視覚が重要である。“スピノサウルス”の吻には内部が連絡した多数の孔があり、ワニ類と同じような機械受容感覚を用いていたと考えられる。“スピノサウルス”は嗅覚よりも機械受容感覚により大きく依存し、イリタトルやバリオニクス亜科ではより嗅覚に依存していた可能性がある。
面白いことに化石記録にはそれを支持するものがある。バリオニクスの腹腔からは半ば消化されかけた魚のウロコとともに、鳥脚類の幼体の骨が見つかっている。またスコミムスの下顎の力学的特性から、スコミムスは小型の陸生動物をも捕食できただろうと示唆されている。さらにアラリペ盆地の翼竜の頸椎にスピノサウルス類の歯冠が埋まっていたことから、この地域のスピノサウルス類が魚以外の獲物を捕食したことが示されている。一方、“スピノサウルス”の歯槽の位置にはOnchopristisとされる魚の脊椎骨の一部が見つかっている。これは間接的な証拠であるとしても、“スピノサウルス”の吻の形態とともに魚類が主な獲物であったことを示唆している。
つまり現在得られている証拠は、バリオニクス亜科とアラリペ盆地のスピノサウルス亜科(イリタトルなど)が陸生動物をも捕食したことを示しており、そのためには嗅覚が重要である。一方“スピノサウルス”が魚類など水生動物を捕食するためには、視覚や機械受容感覚が重要だったと考えられる。
個人的に一番感心したのは、あきらめてはいけないということである。
イリタトルの歯列が同定されていないことは、「盲点」だったのだろう。既に記載され、再記載された標本であっても、アイデア一つで、つまり新しい観点や解釈があれば、新しいデータを得ることができ、論文が書ける。系統解析もやり直すことができる。そこが素晴らしい。
また外鼻孔が保存されたスピノサウルス類は4種しかいないが、外鼻孔周囲の骨の位置関係はそれぞれ異なっているのが面白い。
コメント ( 0 ) | Trackback ( 0 )
ブラジルのスピノサウルス類の再検討とスピノサウルス類の頭蓋の進化 (中)
Sales and Schultz (2017)によるスピノサウルス類の系統解析
イリタトルとアンガトゥラマの関係について、著者らはかなり字数を割いて考察している。イリタトルとアンガトゥラマは、吻以外の頭骨と吻の先端部であり、同じ地層(アラリペ盆地のRomualdo Formation)から発見されたことから、同一種あるいは、ひょっとすると同一個体とさえ考えられたことがある。今回イリタトルの上顎歯列が同定されたことで、最も前方の歯はm3と考えられた。アンガトゥラマの最後の歯もm3なので、この2つの標本はギリギリ重複しており、少なくとも同一個体でないことはわかる。しかし同一種の可能性はあり、そこはまだわからないという。重複しているm3を解析した結果、同種か別種かについて何か情報が得られたのかと期待したが、そこまではわからないようである。おそらくどちらもほとんど同じ形態をしていて、同種とも別種とも言えないのだろう。
またアンガトゥラマの前上顎骨はバリオニクスと同じくらいの大きさなので、ホロタイプの復元頭骨はイリタトルよりも大きいだろうといっている。ホロタイプ自体はそうかもしれないが、種としては成長段階の違いや個体差もありうるし、これだけでは結論できないだろう。
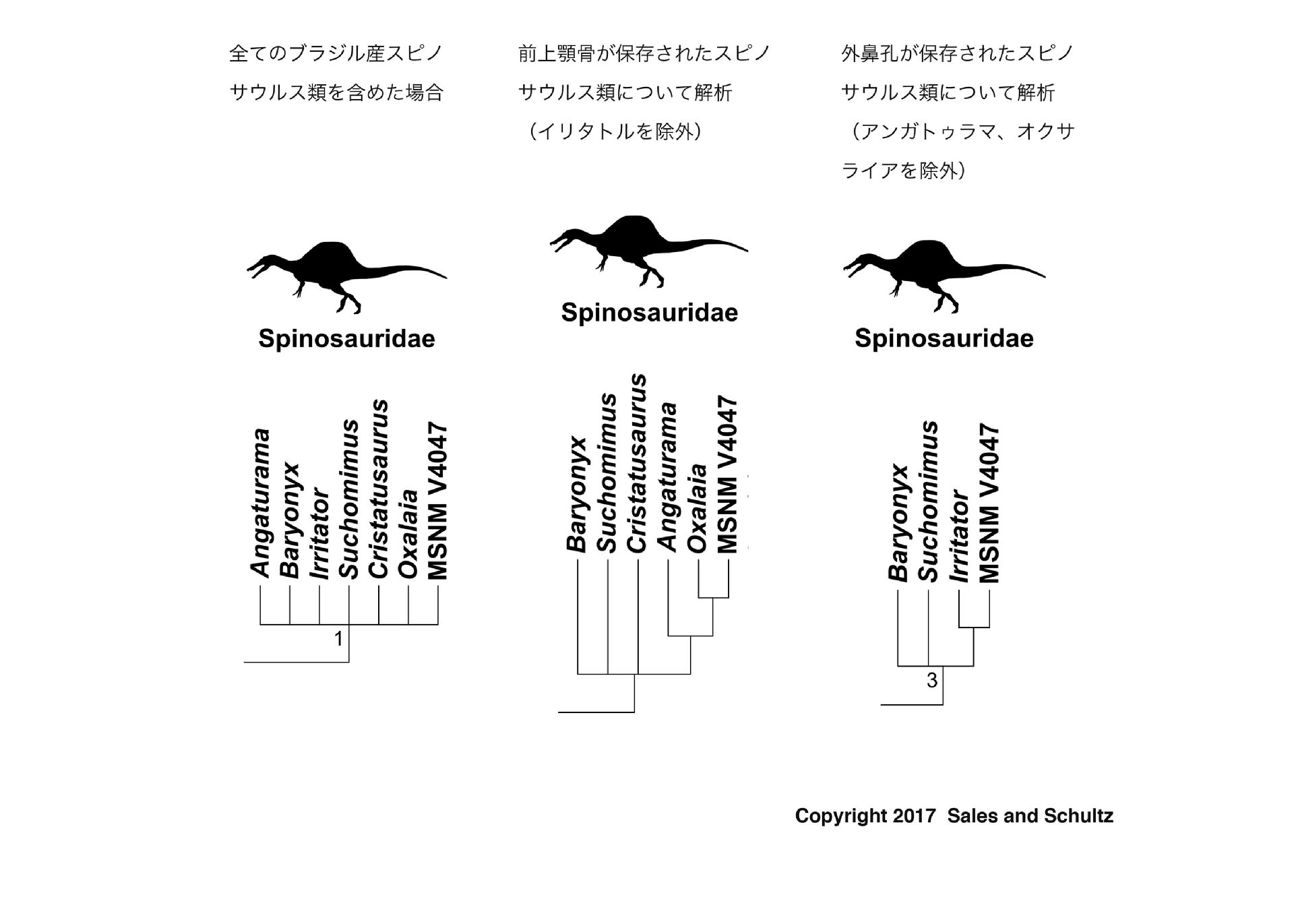
系統解析は、Carrano et al. のテタヌラ類のデータに、今回ブラジル産スピノサウルス類から得られた新しいデータを含めて解析している。頭蓋と歯の情報に注目するため、頭骨がないイクチオヴェナトルやシギルマッササウルスは除外している。まず、すべてのスピノサウルス類を用いて解析すると、スピノサウルス科の部分は、すべてのスピノサウルス類がポリトミー(多分岐)をなしてしまった(図)。
次に前上顎骨が保存されたスピノサウルス類を用いて、つまりイリタトルを除外して解析すると、バリオニクス、スコミムス、クリスタトゥサウルスと”スピノサウルス亜科”のクレードがポリトミーをなし、スピノサウルス亜科の中ではアンガトゥラマ、オクサライアが”スピノサウルス”に対して順次外群をなした。さらに、外鼻孔が保存されたスピノサウルス類を用いて、つまりアンガトゥラマ、オクサライアを除いて解析すると、バリオニクス、スコミムスと”スピノサウルス亜科”がポリトミーとなり、”スピノサウルス亜科”にイリタトルと”スピノサウルス”が含まれた。
前述のようにスピノサウルス科はバリオニクス亜科とスピノサウルス亜科に分けられてきた。バリオニクス亜科は、細かい鋸歯のある、よりカーブした歯冠をもち、前上顎骨の歯隙がはっきりしない。一方スピノサウルス亜科は、鋸歯がないまっすぐな円錐形の歯冠をもち、前上顎骨の一部にはっきりした歯隙がある。また外鼻孔の位置はバリオニクス亜科の方がスピノサウルス亜科よりも前方にある。さらに上顎骨の歯の数が、バリオニクス亜科ではスピノサウルス亜科の2倍近くある。
著者らの2番目と3番目の解析結果では、バリオニクス亜科のメンバーが単系のクレードをなさなかった。実際、多くのバリオニクス亜科の特徴は、原始的な獣脚類の状態と派生的なスピノサウルス亜科の状態の中間段階に相当するものである。例えばスピノサウルス類の鋸歯の進化史を考えれば、獣脚類は最初、大きな鋸歯を持っていたが、次にバリオニクス亜科のような細かい鋸歯となり、ついにはスピノサウルス亜科のように鋸歯を失ったと考えられる。将来、バリオニクス亜科のメンバーはクレードであるスピノサウルス亜科に対して順次外群をなすようになるかもしれない。つまり多系群となるかもしれない。これは、非鳥型獣脚類全体のように原始形質をもつグループにはよくあることである。
今回得られた分岐パターンや頭蓋・歯の特徴の再解釈から、ある程度の進化のシナリオが予想される。獣脚類の前上顎骨の原始状態は、歯が5本以下であった。スピノサウルス科に進化する段階で、歯の数が7本に増加し、terminal rosette が形成された。その後スピノサウルス亜科の前上顎歯列には顕著な歯隙が現れた。おそらく原始状態ではスピノサウルス類の前上顎骨には、バリオニクス亜科とアンガトゥラマにあるような背側正中のとさかがあった。この特徴はその後、スピノサウルス亜科の中のある時点で失われたと考えられる。実際にアンガトゥラマはオクサライアと”スピノサウルス”よりも基盤的であり、オクサライアはどの形質についてもアフリカのスピノサウルス亜科と似ている。
上顎骨歯の大きさの変化パターンがバリオニクス亜科とスピノサウルス亜科で共通していることから、上顎歯列は前方の歯(m1からm4)については、スピノサウルス科の中で相同であると考えられる。しかしバリオニクス亜科の上顎骨歯はスピノサウルス亜科よりも数が多いことから、スピノサウルス類の進化史の中でいくつかの歯は失われたと思われる。つまりm4より後方の歯の相同性は疑わしい。Dal Sasso et al. (2005) は、スピノサウルス亜科では通常、上顎骨歯の間隔が1個の歯槽の大きさと同じくらいであり、このパターンはm4の直後から始まるといっている。つまりこの部分の歯は1個おきに失われた可能性がある。スコミムスの上顎骨歯は22本あり、m4より後方で1個おきに失われたとすれば13本と予測される。”スピノサウルス”MSNM V4047の上顎骨歯は12本あり、イリタトルでは11本である。微妙に一致しないが、”スピノサウルス”とイリタトルでは最後の上顎骨歯が前眼窩窓の前端のレベルにあるのに対して、スコミムスではもっと後方まで延びているという。よってスピノサウルス亜科の歯の減少においては、m4より後方で1個おきに失われたことに加えて、最も後方の歯が失われたことも関与しているかもしれない。
従来の考えと異なり、スピノサウルス科の中で外鼻孔がより前方にあることは、バリオニクス亜科とスピノサウルス亜科のうちのイリタトルに共通している。これらのスピノサウルス類では前上顎骨、上顎骨、鼻骨が外鼻孔を取り囲んでいる。スピノサウルス亜科で外鼻孔がもっと後方に移動する際には、次第に前上顎骨の関与は必要なくなり、ついには”スピノサウルス”のような状態に至ったと思われる。またスピノサウルス科の進化過程では、外鼻孔の大きさが小さくなる傾向がある。この点についてイリタトルは、バリオニクス亜科と”スピノサウルス”の中間の状態を示している。
全般に、アラリペ盆地のスピノサウルス類(イリタトルとアンガトゥラマ)の頭骨は、バリオニクス亜科とアフリカのスピノサウルス亜科の中間的な形質をもっている。一方オクサライアは、よりアフリカのスピノサウルス亜科に近縁である。アンガトゥラマとオクサライアについては、分岐図のように順次外群をなしていると思われる。つまりスピノサウルス亜科は、従来考えられていたよりも形態学的に多様であることがわかってきた。
コメント ( 0 ) | Trackback ( 0 )
| « 前ページ | 次ページ » |