








NHKのテレビ放送で数学講座の1つとして、カオスやフラクタルを解説する講座があった。その中で、野菜のロマネスコは、フラクタルの特徴を備えているとの説明があった。改めてロマネスコの写真を見ると、小花の配列がつくる造形パターンが美しい。また、参考文献1で紹介しているある種のヒマワリの小花がつくる二重らせんパターンが美しい。
ロマネスコとヒマワリの小花の配列は、いずれも植物の葉序パターンの例である。葉序パターンとは、原基と呼ばれる葉のもとになるものの側芽の成長履歴を示すパターンのことである。小花は葉が変形したものであるから、葉序パターンの造形規則が適用される。
多くの植物の茎についた葉の列はらせんを描く。葉はそれぞれ、すぐ下の葉とおおむね一定の角度だけずれたところについている。このずれの角度は多くの種で、黄金角と呼ばれる137.5度に近いことが多い。二重らせんパターンは、原基が、葉ではなく小花に成長するときにはさらにわかりやすい。新しい葉から順に1,2,3,…のように番号をふるとする。右曲がりのらせんの本数が8であるとすれば、左曲がりのらせんの本数は13である。この植物の葉序パターンを(8,13)と表示する。同様に、葉序パターン(13,21)は、右曲がりらせん13本、左曲がりらせん21本をもつ。1本の右曲がりらせん上の小花の番号は、1,14,27,…; 2,15,28,…のように配列される。先に挙げたヒマワリの葉序パターンは、(34,55)と表示される。
(8,13); (13,21)などは、フィボナッチ数列のとなりあった2つのフィボナッチ数である。たとえば21/13=1.615ほどであり、黄金比=1.618034…に近い。黄金比に基づくらせん曲線を描くと、対数らせんに近づいていく。
葉序パターンは、美しい数学的構造をもっているようにみえるので、数理モデルをつくって、フィボナッチ数のペアをもつ葉序パターンが成立するのか否か検証することにした。葉序パターン(13,21)の場合のモデルをつくることにした。開度(小花のずれの角度)は、黄金角の137.5度とした。21/13の比は黄金比に近いので、らせん曲線として対数らせんを前提とするのは自明なようにみえた。しかし、このモデルは、その計算途中で、不適当であることを知ったので、計算を断念することにした。
その理由は、作成したらせん曲線は、番号1の小花から14の小花へと不自然な跳躍をしていることもあるが、大局的にみて、現実の葉序パターンは、らせん曲線に則っているのか否か疑問になったことと、小花と小花との間隔が137.5度の一定値とすることは不自然であることが次第に明確になってきたためである。
改めて参考文献1を読み直してみると、らせん曲線を「フィボナッチらせん」と呼んでいるが、それが対数らせんであるとは言っていない。また、参考文献2では、らせん曲線の用語はなく、「斜列線」の用語で統一している。
葉序パターンの研究は、多くの研究者の興味を引いたとみえて、およそ150年ほどの歴史がある。その間に、研究成果は、深くかつ広くなり、形態学的な解析から理論的枠組みに至るまで、またBZ反応やチューリングの反応・拡散系にまで言及することになり、非線形科学の「パターン形成」という主要な分野を占めることになったようである。
参考文献1を見ていると、数理モデルによって現実の植物の葉序パターンをシミュレーションした例が見当たらない。その代わりと言うか、植物の生育とは関係のない物質を用いて物理実験を行い、パターン(13,21)のような「フィボナッチらせん」を実現できたとする研究成果を二例紹介している。
その一例を引用すると、「(フランスの物理学者は、)油の膜でおおった円盤に、磁気を帯びた液体の小さなしずくを落とした。しずくは油の層に浮いた。装置は磁場の中にあり、磁場によって磁気を帯びたしずくが分極化し、斥けあう。彼らは、水平の磁場も生じさせた。これは円盤の中心よりもヘリで強い。このためしずくはヘリのほうに引っ張られる。したがってしずくは、1つ1つ落ちるにつれて、斥けあいながら円盤のヘリに押しやられる。これは、分裂組織の頂端で新たな原基が形成される様子におよそ類似していると彼らは論じた。しずくは、十分なペースで加えられると外に移動して、(1,2)から(13,21)までさまざまなフィボナッチ二重らせんパターンを形づくる。それぞれのしずくは前のしずくから、137.5度に近い角度だけずれる。」
参考文献1は、この実験から得られたフィボナッチ・パターンの図を提示している。そのパターン図は、二重らせんが各々のしずくを通過する見事なパターンを示している。この図を見ていると、数理モデルをつくってこのパターンをシミュレートすることは容易なようにみえる。しかし、実際にはフィボナッチらせんのシミュレーションは容易ではないようだ。
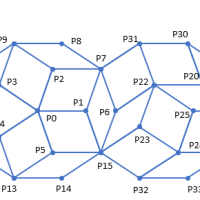
ある数学者は、らせんに沿って連続的に置かれた原基を、むだな空間をあまり残さずに最適な仕方で詰め込むようなモデルを用いて、開度が137.5度とわずかに違うらせん葉序を生成した。すると、この角度が1/10度しか違っていなくても、パターンが大きくなるにつれ要素の密集度はかなりゆるくなり、むだが多くなった。
「生体が(137.5度という)これほどの正確さで組織化できるとは考えられない。実際、らせん葉序をなす葉の開度は植物によって、また同じ植物でも葉によって相当異なる。オナモミについておこなわれたある分析では、角は124度からおよそ150度にまで及ぶことがわかった。そうすると、葉序のフィボナッチ・パターンは、密集効果では説明しつくせない。フィボナッチらせんのペアは、数学的には、黄金角から少しずれがあるだけで崩れてしまうが、現実の植物では、アバウトなところで充分通用する。そこで、葉序のメカニズムは、ほかの基準に支配されているが、たまたま葉の配置が本当に最適な詰め方に近くなるようになっていることを示唆する。」
こうなると、二重らせん構造の葉序パターンをもつ植物のすべてが、フィボナッチ数のペアで表示できるような「フィボナッチらせん」になっているのかどうかさえ疑問になってくる。葉序パターンの裏の顔として上記のような数学的構造が隠れていることは否定できないが、該当する属性をもつすべての植物が周知の数学的構造を厳密に守っているのか否かとなると、相当怪しい。
参考文献2は、2~5個のパラメータを指定するような葉序パターン生成機構の抽象モデルを紹介している。これらパラメータは、原基同士の反発的な作用を数式化するもの、葉間期に関するもの、などを含む。「これらのモデルでは、すでに存在する葉原基は新たな原基の形成を妨げる阻害作用を発し、周囲に阻害場を作り出していること、その影響は葉原基からの距離に応じて減衰すること、新たな葉原基はシュート頂分裂組織周縁部でこの阻害の影響の及ばないところ(影響の充分小さいところ)に形成されることが仮定されている。」葉間期とは、葉の発生の時間間隔のことである。
ここで、またシドニー・ブレナーの言葉: 「数学は完璧を目指す学問。物理学は最適を目指す学問。生物学は、進化があるため、満足できる答えを目指す学問だ」を引用したくなる。
参考文献2で紹介する抽象モデルによるシミュレーション解析は、形態学的な解析から得られるデータに近づけることが可能なようだ。葉序パターン研究の進化は、2023年4月2日付のブログ「なぜ自然は数学の言葉で書かれているのか」という問題に答える一つの事例であると考える。
参考文献
ボール著「かたち 自然が創り出す美しいパターン」(早川書房)
米倉崇光ほか著「葉序の形態特徴量とパターン生成機構との関係」(PLANT MORPHOLOGY vol 33 pp59~66)
ロマネスコとヒマワリの小花の配列は、いずれも植物の葉序パターンの例である。葉序パターンとは、原基と呼ばれる葉のもとになるものの側芽の成長履歴を示すパターンのことである。小花は葉が変形したものであるから、葉序パターンの造形規則が適用される。
多くの植物の茎についた葉の列はらせんを描く。葉はそれぞれ、すぐ下の葉とおおむね一定の角度だけずれたところについている。このずれの角度は多くの種で、黄金角と呼ばれる137.5度に近いことが多い。二重らせんパターンは、原基が、葉ではなく小花に成長するときにはさらにわかりやすい。新しい葉から順に1,2,3,…のように番号をふるとする。右曲がりのらせんの本数が8であるとすれば、左曲がりのらせんの本数は13である。この植物の葉序パターンを(8,13)と表示する。同様に、葉序パターン(13,21)は、右曲がりらせん13本、左曲がりらせん21本をもつ。1本の右曲がりらせん上の小花の番号は、1,14,27,…; 2,15,28,…のように配列される。先に挙げたヒマワリの葉序パターンは、(34,55)と表示される。
(8,13); (13,21)などは、フィボナッチ数列のとなりあった2つのフィボナッチ数である。たとえば21/13=1.615ほどであり、黄金比=1.618034…に近い。黄金比に基づくらせん曲線を描くと、対数らせんに近づいていく。
葉序パターンは、美しい数学的構造をもっているようにみえるので、数理モデルをつくって、フィボナッチ数のペアをもつ葉序パターンが成立するのか否か検証することにした。葉序パターン(13,21)の場合のモデルをつくることにした。開度(小花のずれの角度)は、黄金角の137.5度とした。21/13の比は黄金比に近いので、らせん曲線として対数らせんを前提とするのは自明なようにみえた。しかし、このモデルは、その計算途中で、不適当であることを知ったので、計算を断念することにした。
その理由は、作成したらせん曲線は、番号1の小花から14の小花へと不自然な跳躍をしていることもあるが、大局的にみて、現実の葉序パターンは、らせん曲線に則っているのか否か疑問になったことと、小花と小花との間隔が137.5度の一定値とすることは不自然であることが次第に明確になってきたためである。
改めて参考文献1を読み直してみると、らせん曲線を「フィボナッチらせん」と呼んでいるが、それが対数らせんであるとは言っていない。また、参考文献2では、らせん曲線の用語はなく、「斜列線」の用語で統一している。
葉序パターンの研究は、多くの研究者の興味を引いたとみえて、およそ150年ほどの歴史がある。その間に、研究成果は、深くかつ広くなり、形態学的な解析から理論的枠組みに至るまで、またBZ反応やチューリングの反応・拡散系にまで言及することになり、非線形科学の「パターン形成」という主要な分野を占めることになったようである。
参考文献1を見ていると、数理モデルによって現実の植物の葉序パターンをシミュレーションした例が見当たらない。その代わりと言うか、植物の生育とは関係のない物質を用いて物理実験を行い、パターン(13,21)のような「フィボナッチらせん」を実現できたとする研究成果を二例紹介している。
その一例を引用すると、「(フランスの物理学者は、)油の膜でおおった円盤に、磁気を帯びた液体の小さなしずくを落とした。しずくは油の層に浮いた。装置は磁場の中にあり、磁場によって磁気を帯びたしずくが分極化し、斥けあう。彼らは、水平の磁場も生じさせた。これは円盤の中心よりもヘリで強い。このためしずくはヘリのほうに引っ張られる。したがってしずくは、1つ1つ落ちるにつれて、斥けあいながら円盤のヘリに押しやられる。これは、分裂組織の頂端で新たな原基が形成される様子におよそ類似していると彼らは論じた。しずくは、十分なペースで加えられると外に移動して、(1,2)から(13,21)までさまざまなフィボナッチ二重らせんパターンを形づくる。それぞれのしずくは前のしずくから、137.5度に近い角度だけずれる。」
参考文献1は、この実験から得られたフィボナッチ・パターンの図を提示している。そのパターン図は、二重らせんが各々のしずくを通過する見事なパターンを示している。この図を見ていると、数理モデルをつくってこのパターンをシミュレートすることは容易なようにみえる。しかし、実際にはフィボナッチらせんのシミュレーションは容易ではないようだ。
ある数学者は、らせんに沿って連続的に置かれた原基を、むだな空間をあまり残さずに最適な仕方で詰め込むようなモデルを用いて、開度が137.5度とわずかに違うらせん葉序を生成した。すると、この角度が1/10度しか違っていなくても、パターンが大きくなるにつれ要素の密集度はかなりゆるくなり、むだが多くなった。
「生体が(137.5度という)これほどの正確さで組織化できるとは考えられない。実際、らせん葉序をなす葉の開度は植物によって、また同じ植物でも葉によって相当異なる。オナモミについておこなわれたある分析では、角は124度からおよそ150度にまで及ぶことがわかった。そうすると、葉序のフィボナッチ・パターンは、密集効果では説明しつくせない。フィボナッチらせんのペアは、数学的には、黄金角から少しずれがあるだけで崩れてしまうが、現実の植物では、アバウトなところで充分通用する。そこで、葉序のメカニズムは、ほかの基準に支配されているが、たまたま葉の配置が本当に最適な詰め方に近くなるようになっていることを示唆する。」
こうなると、二重らせん構造の葉序パターンをもつ植物のすべてが、フィボナッチ数のペアで表示できるような「フィボナッチらせん」になっているのかどうかさえ疑問になってくる。葉序パターンの裏の顔として上記のような数学的構造が隠れていることは否定できないが、該当する属性をもつすべての植物が周知の数学的構造を厳密に守っているのか否かとなると、相当怪しい。
参考文献2は、2~5個のパラメータを指定するような葉序パターン生成機構の抽象モデルを紹介している。これらパラメータは、原基同士の反発的な作用を数式化するもの、葉間期に関するもの、などを含む。「これらのモデルでは、すでに存在する葉原基は新たな原基の形成を妨げる阻害作用を発し、周囲に阻害場を作り出していること、その影響は葉原基からの距離に応じて減衰すること、新たな葉原基はシュート頂分裂組織周縁部でこの阻害の影響の及ばないところ(影響の充分小さいところ)に形成されることが仮定されている。」葉間期とは、葉の発生の時間間隔のことである。
ここで、またシドニー・ブレナーの言葉: 「数学は完璧を目指す学問。物理学は最適を目指す学問。生物学は、進化があるため、満足できる答えを目指す学問だ」を引用したくなる。
参考文献2で紹介する抽象モデルによるシミュレーション解析は、形態学的な解析から得られるデータに近づけることが可能なようだ。葉序パターン研究の進化は、2023年4月2日付のブログ「なぜ自然は数学の言葉で書かれているのか」という問題に答える一つの事例であると考える。
参考文献
ボール著「かたち 自然が創り出す美しいパターン」(早川書房)
米倉崇光ほか著「葉序の形態特徴量とパターン生成機構との関係」(PLANT MORPHOLOGY vol 33 pp59~66)
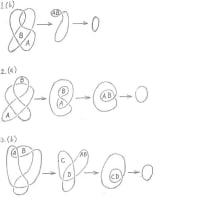


※コメント投稿者のブログIDはブログ作成者のみに通知されます