中国(および東アジア)の蝶Butterflie of China(and Eastern Asia)中国(和东亚)的蝴蝶
クロキマダラヒカゲ 田园荫眼蝶 キマダラヒカゲ属 Neope荫眼蝶 【タテハチョウ科Nymphalidae蛱蝶科】
Junzo Aoyama
【Photo office萌葱】
はじめに
中国をフィールドにして35年が経つ(この3年間は新型ウイルスとかで日本に閉じ込められているが)。1998年に400頁余に及ぶ「中国のチョウ」を刊行してからも、倍以上の24年が経った。その間、何冊かの自主刊行本(ミヤマシロチョウ、ベニシジミなど)を作成したが、筆者の資金力では数冊の製本が精一杯で、その後は途絶えてしまった。
作品自体は、ほかにも数多く書き溜めてはいる。しかし書籍化することは困難である。ということで、このままでは写真や資料を死蔵したままになってしまう可能性が高い。そこで、これまでに書き溜めて来た「未完成作品」をクオリティには目をつぶって(廉価な印刷製本で)世に問うことにした。
一部は自主刊行本として発表済みの作品のリニューアルであり、また「中国のチョウ」(東海大学出版会1998)の記事の再掲も行った。構成上も内容的にも中途半端なスタイルであり、多くの内容が重複しているが、敢えてそのままの形で組むことにした。本来なら、到底商品として成り立つものではないと思うが、筆者は今後も改めて「中国と日本の蝶」の探索に挑戦したいという想いがあり、それを実現するためのカンパだと解釈して頂ければ幸いである(写真のクオリティに関しては付録にオリジナル原稿を収納したCDを添えることでカバーが出来ると思う)。
蝶の研究者や愛好家やコレクターの知識量は凄まじいものがある。著者などは、文字通り足元にも及ばない、と自覚している。ただ、著者は30年間以上に亘って、自分の脚で各地を歩き、自分の眼で生きた蝶たちの姿に接し、自分の頭でその実態再現を構築してきた、という自負もある。上記した人々(研究者や愛好家)とは別の次元からの、対象の把握が為されていると思っている。
体系的にではなく俯瞰的に、細部の徹底追及ではなくランダムな拾い読みで、斜め横辺りの視点から、様々な問題提起を行っていこうと思っている。それはそれで、意味のある事であろう。
とは言っても、なにしろ、ここ数年はフィールドからも遠ざかっているし、「中国のチョウ」上梓以降は、ゲニタリアの検鏡も出来ないでいる。古い知識に頼った、ほとんど感覚的ともいえる分析ばかりである。著者の指摘が、お門違いだったり、幼稚に過ぎるという批判を受けるだろうことは、承知している。
本を購入して頂いた、すなわちお金をとっていながらこのようなお願いをすることは厚かましいこと限りないのだが、 著者の呈した様々な疑問(問題提起)などについて、忌憚の無いご意見を頂ければ有難い。
著者の呈した問題提起を、読者の皆さんと一緒に考えていくことが出来ればと望んでいる。一応「マガジン」という性格を利用して、意見をくだされば、次号以降に誌上で議論を展開していくことも出来ると思う。
どうか、よろしく。
*作品によっては「青山潤三遺稿集」となっています。その時点における将来(正式作品発表時)を考慮しての準備処置であることを御理解ください。また、著者として、若い友人の李芳Li Fang(Monica Lee)/Lewis Crannitchの名が加えられている場合がありますが、撮影・執筆はほぼ全て青山によるものです(翻訳の一部を両名が担当)。両名のクレジットが為されているものについては将来を考え、それを付したままにしておきます。
*オリジナル原稿と原版写真を収納したCDを添付しました。原版写真の個人的利用は自由に行ってください。商業的に使われる場合は、その旨を著者へ報告いただき、収益の一部(各自のご判断に任せます)を納入ください。
/////////////////////
週刊:中国(および東アジア)の蝶 【2022.1.8】 『クロキマダラヒカゲNeope agrestis(キマダラヒカゲ属 Neope)』は、主に次の原典を元に構成しています。This book is based on the following original text. 本书根据以下原文改编。
●作成途上の単行本「中国の蝶」のNeope属の項の抜粋 2020.7.4脱稿(CD版「青山潤三作品集」収納)訳文追加
●「日本固有種ヤマキマダラヒカゲ・サトキマダラヒカゲの大陸産対応種について」
(ブログ青山潤三の世界あや子版2013.5.24)の再録(写真を除く記事全文、*は本書での注と改変)
●「中国のチョウ(東海大学出版会)1998」の再録(写真を除く記事全文)
ジャノメチョウ族Satyrini/ヒカゲチョウ亜族Lethina/キマダラヒカゲ属Neope荫眼蝶属
筆者が撮影したことのある中国大陸産(および日本産2種)
Neope yama 丝链荫眼蝶 オオキマダラヒカゲ
Neope muirheadii 蒙链荫眼蝶 ウラキマダラヒカゲ
Neope simulans 拟网荫眼蝶 ウラナミキマダラヒカゲ
Neope armandii 阿芒荫眼蝶 シロキマダラヒカゲ
Neope ramosa 大斑荫眼蝶(暫定分類) タイリクアリサンキマダラヒカゲ(台湾産を含むpulahaとの関係は未詳)
Neope pulahoides 黑斑荫眼蝶(暫定分類) オオアリサンキマダラヒカゲ(台湾産を含むpulahaとの関係は未詳)
Neope oberthueri 奥荫眼蝶 スミイロキマダラヒカゲ
Neope goschkevitchii 日本荫眼蝶 サトキマダラヒカゲ(日本列島固有種)
Neope agrestis 田园荫眼蝶 クロキマダラヒカゲ
Neope niphonica 山荫眼蝶 ヤマキマダラヒカゲ(日本列島固有種)
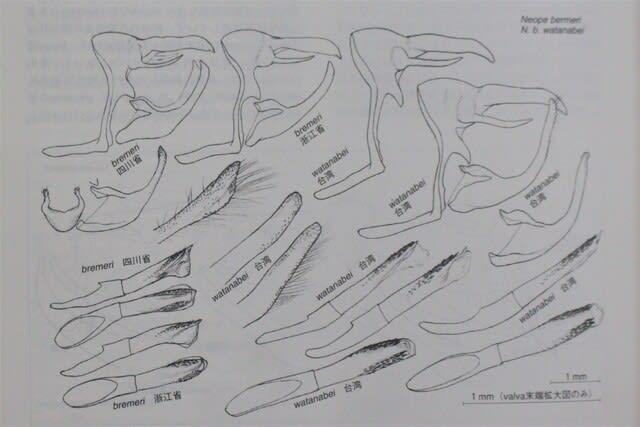
Neope buremeri 布莱荫眼蝶 アカキマダラヒカゲ(台湾産タイワンキマダラヒカゲの原名亜種)
Miller(1968)などによる古くからの分類でも、DNA解析に基づいた新しい分類体系でも、キマダラヒカゲ属は常にヒカゲチョウ族(あるいはジャノメチョウ族ヒカゲチョウ亜族)の一員として扱われてきた。筆者もこの処遇を概ね支持するが、「必ず翅を閉じて止まる」という、他のヒカゲチョウ亜族の各種と異なる独自の性質を持つことは、非常に気になる点である。ただし、キマダラヒカゲ属のうち、Neope yama において♂ゲニタリアの構造がヒカゲチョウ属の一グループ(独立属と見做した場合のZophoessa属)との共通点を有している事は、本種がNeope属とLethe属(Zophoessa属)との橋渡し的存在であることを示しているように思われ、生態上の特徴は二次的な現象なのかも知れない。
Neope agrestisとNeope oberthueriiは、Neope属全体として見た場合、ごく狭い地域にのみ分布する希少種だが、梅里雪山周辺では最もポピュラーな蝶の一つである。15年間に亘りこの地で調査・撮影を続けた著者にとっては、最も身近な対象であるため、本書ではこの2種を中心に紹介していく。
Neope属は、日本でもよく似た2種(ヤマキマダラヒカゲとサトキマダラヒカゲ)がセットになって、大局的に見れば(原則サトキマダラヒカゲのみ分布する低標高地・温暖地を除く)分布圏のほぼ全域で混在している。同様に大陸部に於いても、属の分布域全体(「東アジア」地域、ただし朝鮮半島周辺を除き、ヒマラヤ地方を含む)で2つの種*がセットで分布しているように思える。それらの“2種”が日本産2種との間に、どのような系統的反映を示すかという判断はさておき、それぞれの地における近縁2種(あるいは任意の分類群)の関係性は、日本産2種間の関係性と同一方向を示している、という実態は確かなようである。
ごく大雑把に捉えれば、agrestis-niphonica‐bremeri(台湾産watanabeiを含む)と、oberthueri‐goschkevitchii
‐pulaha(complex)の流れを読み取れる。系統的な直接の繋がりの是非(ことに後者の組み合わせでは積極的な支持要素が見つからない)はともかく、雄交尾器、斑紋様式、雄の性標の状態、等々の比較の上では、ある程度の纏まりを持つようにも思えるし、かつ非常に複雑で混沌としている部分もある。DNAの解析も含め、今後の検証を待たねばならない。
*ほかに、やや異なる系統のシロキマダラヒカゲ近縁種群やウラキマダラヒカゲ(この両種は台湾にも分布)、ウラナミキマダラヒカゲ、オオキマダラヒカゲなども幾つかの地域で同所分布することを確認している。Neope 属としては他にも幾つかの種(全て中国西南部‐インドシナ半島北部‐ヒマラヤ地方)が記載されているが、筆者は未確認のため、詳細については言及し得ない。明らかな独立種(ただしarmandiとの関係については検討余地が残る)であろうbhadraなど数種を除いては、本書でとり上げた各種に収斂されるものと思われる(その場合の模式分類群の特定については未詳)。
**和名は、原則として「中国のチョウ」で取り上げた名を使用した。外観の印象に照らし合わせて、必ずしも適切で無いものも含まれると思うが、そのままにしておく。
/////////////////////////
Neope agrestis 田园荫眼蝶 クロキマダラヒカゲ 《bremeri種群》
写真(省略)上:雲南省梅里雪山明永2017.6.15alt.2500m/下左:同2009.6.12 alt.3600m/下右:維西~巨旬2010.5.18 alt.2500m
>四川省の大渡河以西から雲南省西北部を経てミャンマー北部に至る標高2000m以上の高地帯に分布し、次種と混棲。長江中~下流域低地帯に分布するNeope bremeri(=台湾産タイワンキマダラヒカゲ原名亜種)の代置的存在で、同様に後翅裏面基部近くの三丸紋の下側の一つが顕著にずれるが、亜外縁沿いの眼状紋は余り離れずに並ぶ。眼状紋中心の白点を欠く。 おそらく年1化。4月から7月まで新鮮個体が見られる。
>Coexists with next species Neope oberthueri in the mountainous areas of southwestern China. A substitute species of Neope bremeri (distributed in the lowlands of middle to lower reaches of 長江), similar to bremeri, one of lower sides of 3 crest near base of back side of thind wing is significantly displaced. However, top 3 of the eye-spots along outer edge are not apart. Lacks white spot in the center of eyespot. One brood from April to July.
>在中国西南山区与下一个物种共存。它是分布在长江中下游低地的布莱荫眼蝶(=台湾荫眼蝶原名亚种)的替代种。但, 沿外边缘的成排眼点, 上三个也排成一排。它缺少眼点中心的白点。年发生一次(4‐7月)。
Neope oberthueri 奥荫眼蝶 スミイロキマダラヒカゲ 《pulaha種群》
写真(省略)上:雲南省梅里雪山明永2017.6.15 alt.2300m/下左:同/下右:梅里雪山西当~雨崩2009.6.11 alt.3300m
>中国西南部山岳地帯(雲南省西北部~ミャンマー北部)に分布しNeope agrestisと混棲する。色彩・斑紋の個体差が著しいが、後翅裏面基部の3丸紋が離れずに並び、前表翅脈沿いが暗色(同所分布するsimulansと区別不可)であること等は共通。後翅裏面外縁沿いの上から2つ目の眼状紋は前種同様余り内側に寄らない。翅裏は薄紫色を帯びることが多い。4月~7月に出現するが、おそらく年1化。
>Distributed in mountain areas of southwestern China and coexists with Neope agrestis. Significant individual differences in color and mottled pattern. 3 round patterns at base of back face of hind wings are lined up straight. vein of upper face are dark (indistinguishable from simulans). 2nd eye-spot from top along outer edge of back face of hind wing don't move inward same as agrestis. Back face is often light purple. One brood April to July.
>分布于中国西南部山区,与奥荫眼蝶共存。 颜色和斑纹个体差异显着,但共同点是后翅背面基部的3个圆形花纹排列成直线,前翅脉较暗(与拟象无区别)。沿后翅背面外缘自上而下的第2个眼点不向内移动,与奥荫眼蝶相同。 翅膀背面常呈淡紫色。年发生一次(4‐7月)。
Neope bremeri 布莱荫眼蝶 アカキマダラヒカゲ(タイワンキマダラヒカゲ原名亜種)
写真(省略)浙江省杭州市alt.10m 1989.3.31(春型♂)/四川青城山alt.900m 1989.8.4(夏型♂)/浙江省清涼峰alt.400m 2018.8.14(夏型♂)/浙江省杭州市alt.10m 1989.3.31(春型♀)
>長江中~下流域に広く分布し、河口近くの杭州では都心にも棲息。成都西郊の低山地にも分布しているが、それ以西の西南部高地帯ではNeope agrestisに置き換わる。後翅裏面基部の丸紋、亜外縁沿いの眼状紋ともに2つ目と3つ目が顕著にずれる(agrestisとは前者で共通し後者で相違する)。雄翅表に特殊鱗密集域がある(上下には分離しない)。年二化。春型(次頁上右も)と夏型(次頁左下も)で外観差が著しい。
>Widely distributed in middle to lower reaches of 長江, also inhabits city center in 杭州 near mouth of 長江. Also distributed in low mountains west of 成都, but it replaces to Neope agrestis in high-altitude mountain area on west. Both, rows of round crests at the base of back of hind wings and rows of eye-spots along sub-outer edge are second and third are significantly off (former is same to agrestis and latter is different). Male wing surface have special scaled area (not separated above and below). Significant difference in appearance between the spring type (also on the upper right of the next page) and the summer type (also on the lower left of the next page).
> 广泛分布于长江中下游(包括近长江口杭州市中心)、在成都西郊的低山也有分布,但它取代它西部高地的 Neope agrestis。后翅背面基部圆冠,沿外缘的一排眼点,两个都,第二个和第三个显着交替(与agrestis比较,前者同,后者不同)。牡机翼表面覆盖着特殊的鳞片区域(上下不分开)。春季型和夏季型在外观上存在显着差异。
Neope ramosa大斑荫眼蝶₋pulahoides黑斑荫眼蝶₋pulaha complex
写真(省略)四川省西嶺雪山alt.1100m 1991.4.9/四川省青城山alt.900m 1990.4.8、同・1989.4.5(参考Neope bremeri春型)/中段と下段左(参考Neope bremeri夏型)と中央:浙江省西天目山alt.1300m 2018.7.8、下段右:四川省西嶺雪山alt.1500m 2009.8.5
>四川省大渡江以西のNeope agrestisとNeope oberturi、長江下流域のNeope bremeriは種名を特定したが、その他の同定についてはギブアップ。参考としてbremeri春型(上右)と夏型(下左)も紹介しておく。上中央は性標欠如、右(bremeri)は有り、他は未確認。西天目山の個体はbremeriを含め全て同一地点・日時の撮影。
>Identify species names for N.agrestis and N.oberturi west of 四川大渡河, and N.bremeri lower reaches of長江, but give up for other identifications. For reference, also introduce bremeri Spring form (upper right) and Summer form (lower left). Upper center lacks special scaled area, right (=bremeri) present, others are unconfirmed. All individuals of 西天目山(including bremeri) photographed at same location and at same date and time.
>对四川大渡河以西的Neope agrestis、Neope oberturi、长江下游的Neope bremeri 进行种名鉴定、放弃其他鉴定。作为参考,还介绍了 bremeri夏季型(右上)和夏季型(左下)。上中特殊的鳞片无,右(=bremeri)有,其他未确认。 西天目山(包括bremeri)都是在同一地点同一日时拍摄。
Neope pulahoides 黑斑荫眼蝶 オオアリサンキマダラヒカゲ (附:ユンナンクロキマダラヒカゲ)
写真(省略)越南(ベトナムVietnam)沙巴(Sa-pa)2009.3.13 /越南沙巴alt.1500m 2009.3.10/越南沙巴alt.1400m 2011.7.25/ 雲南省金平alt.2000m 1995.4.8(ユンナンクロキマダラヒカゲ)
>春夏とも大型で、翅型が鋭角なことなどNeope armandiに類似(目を窄めて見ると後翅裏基方寄りのarmandiの白帯と同じ位置にぼんやりと白帯が走るようにも思える)。ここでは暫定的にNeope pulahoidesとしておく。下右は雲南省最南部のベトナム国境山地で4月上旬に撮影(やや汚損した個体が多く、同時期同所に見られるarmandiは新鮮)。Neope agrestisやNeope bremeriに対応すると思われるが、翅表地色が著しく黒く(それについてはNeope obertureiとの関連も考えねばならない)、♂交尾器はNeope pulahaなどにも似る。
>As a whole, similar to Neope armandi in that both spring and summer form are large and acute-angled wing. When squeeze eyes, it seems that the white belt runs vaguely at the same position as the white belt of armandi near the back of the hind wings. Here, it is tentatively indentify to as Neope pulahoides. Bottom right, photographing in early April in the Vietnam border mountainous area in the southernmost part of Yunnan (many individuals are slightly soiled, and armandii are fresh, that I checked at same time same place). It seems to correspond to Neope agrestis and Neope bremeri, but wing surface color is remarkably black (also need consider relationship with Neope oberturei) and ♂ genitalia is similar to Neope pulaha.
>作为一个整体与 Neope armandi 相似、春夏两季都很大、有一个锐角的翅膀形状(挤眼一看,白带似乎在后翅后部附近的armandii白带的同一位置依稀可见)。我暂时称 Neope pulahoides。右下图是4月初在云南省最南端的越南边境山区拍摄的(很多都是轻微的脏,同时在那里看到的Neope armandi是新鲜的)。它似乎对应于 Neope agrestis 和 Neope bremeri,但翼面颜色显着黑色(也必须考虑与 Neope oberturei 的关系),♂生殖器与Neope pulaha 相似。
Neope armandii 阿芒荫眼蝶 シロキマダラヒカゲ
写真(省略)四川省西嶺雪山alt.2000m 2011.7.11/雲南省金平alt.2000m 1995 .4.11/四川省峨眉山alt.1700m 1990.6.1/雲南省金平alt.2000m 1995 .4.11
>後翅表が白~オレンジ色を帯び、後翅裏基方寄りに明確な白帯が斜めに走る(この特徴がさらに強調されたのがNeope bhadraだが、系統的関連については未詳)。四川省産は後翅表が白色、雲南省金平産と広西壮族自治区興堂山産(2007.4.30図示なし)は橙褐色を帯びる(裏面にも出現)。雄交尾器はプロポーションがやや異なる以外は他の各種と共通(全体的に見てNeope agrestisが最も寸詰まりで本種が最も前後に長い)。
> Inside of hind wings white or pale orange, and outside clear white belt runs diagonally toward base to upper edge (this feature is further emphasized by Neope bhadra, but systematic relationship is unknown). Inside of hind wings color 四川 is white, 雲南and広西 (not shown this book, April 30, 2007) are orange-brown (also it appear on outside). The male genitalia is common to all other species except that the proportions are slightly different (overall, Neope agrestis is most compact and Neope armandii is longest in front to back).
>后翅正面白色和淡橙色,背面有一条清晰的白带斜向基部至上缘(Neope bhadra 进一步强调了这一特征,但系统关系未知)。后翅内侧颜色四川为白色,云南和広西(本书未显示,2007年4月30日)为橙棕色(也出现在外侧)。雄性生殖器与所有其他物种相同,只是总体比例有不同(总体而言,Neope agrestis 最紧凑,而 Neope armandii 从前到后最长)。
Neope muirheadii 蒙链荫眼蝶 ウラキマダラヒカゲ
写真(省略)広東省紹関市翁源県2015.3.29/越南沙巴2009.3.11/広東省紹関市翁源県2013.6.12
>中国南部の低地帯で最も普通に見られるNeope属の種。翅表は一様に褐色(亜外縁に眼状紋列がある)。裏面に白帯があり、春型では主に前翅に細く現れるだけ(一見bremeriなどに似た印象)だが、夏型では後翅を含みより太く明瞭に出現する。♂交尾器はuncus先端が二分する。大型種で雌雄は類似し♀はやや翅色が淡く丸みを帯びる傾向がある。生態的地位は、日本のサトキマダラヒカゲにほぼ一致する。
>Most common species of the genus Neope in lowlands of southern China. Wing surface is uniformly brown (with eye-spots line along sub-outer edge). Outside have a white belt, Spring form barely appearance thinly and only front wing (at first glance same as bremeri). Summer type it appears thicker and clearer including hind wings. Uncus of male genitalia is bisected. large species, male and female are similar(female wings slightly round) . Ecological niche is almost same as Japan's Neope goschkevitchii.
>中国南方低地最常见的尼奥佩属种。机翼表面呈均匀的棕色(亜外缘有眼点排)。背面有一条白色的带子,春季型只出现在前翼上薄薄的(乍一看,很像布雷梅里)夏季型前翼后翼都出现厚实清晰白帯。体型较大,雌雄相近,雌翅略和圆形。雄 uncus 的尖端被一分为二。生态位几乎与日本的N.goschkevitchii相同。
a type of Neope muirheadii ? 蒙链荫眼蝶的一型?
写真(省略)/広西壮族自治区興安県猫児山南麓2005.4.23/同/広西壮族自治区龍勝県猫児山北麓2009.4.22
>おそらくNeope muirheadiiの一型(北部産春型?)と思われるが、眼状紋が著しく小さくなるなど、極めて顕著で、かつ地域的に安定的な特徴を示すことから、別途に紹介しておく。湖南省との境に近い広西壮族自治区猫児山北麓と南麓の、渓流沿いの鬱閉した竹林や照葉樹林内に棲息する。
>Probably a type of Neope muirheadii (spring form in northern area in China?), but it is extremely remarkable shows regional stable characteristics (such as extremely small eye-spots), so I will introduce it separately. Lives in bamboo forest beside coppice, at north and south face of 猫児山(広西壮族自治区near border of 湖南省).
>可能是Neope muirheadii的一型(中国北方地区的春季形态?),但它非常显着,表现出区域稳定的特征(如极小的眼点),所以我将单独介绍。 生活在猫児山南北面的灌木丛旁的竹林中(広西壮族自治区湖南省边境附近)。
Neope simulans 拟网纹荫眼蝶 ウラナミキマダラヒカゲ
写真(省略)四川省ミニャコンカ海螺溝alt.1800m 1989.5.3/雲南省梅里雪山明永alt.2300m(屋内侵入個体)2017.6.15
>四川省最高峰ミニャコンカ(7556m)山麓の海螺溝氷河下方と雲南省最高峰梅里雪山(6740m)山麓の明永氷河下方で撮影した。梅里雪山では明永村の入山管理事務所の室内の床に多数のNeope agurestisに交じって死骸が散乱していた。翅表の色彩・斑紋は、同所に分布するもう一つの種Neope oberutueriと区別できない。後翅裏中室端(第4室基部)に明茶褐色の特殊鱗粉を密生する。
>Both pictures were taken at below glacier of Minya-konka (7556m), the highest peak in Sichuan, and Meili Snow Mountain (6740m), the highest peak in Yunnan. In Meili Snow Mountains, dead bodies were scattered on the floor of mountain management office, mixed with many Neope agurestis. Color and mottle on the wing surface are indistinguishable from another species, Neope oberutueri, which is distributed in same place. Light brown special scales are densely grown on edge of middle chamber behind hind wings (base of the 4th chamber).
>两张照片均拍摄于四川最高峰貢夏山(7556m)和云南最高峰梅里雪山(6740m)冰川下方。 在梅里雪山,山管处的地板上散落着尸体(混杂着许多Neope argestis)。 翅膀表面的颜色和斑驳与分布在同一地点的另一个物种 Neope oberutueri 无法区分。 后翅后中室边缘(第4室基部)密生淡褐色特殊鳞片。
Neope yama 丝链荫眼蝶 大黄斑日陰蝶
写真(省略)四川省峨眉山alt. 1700m 1990.6.1
>Neope属中の最大型種。♂交尾器のpennisの形状が、前後に短く、左右に幅広く、背方に強く屈曲して、その腹方に広くperi-vesical areaを生じることなど(一般的にはLethe属に含められる)Zophoessa属各種のそれと共通する。両者(Zophoessa属各種とNeope yama)による祖先的形質の共有と見る事が出来るかも知れない。
>The largest species in genus Neope. Pallus of male genitalia is short, wide in the left and right, and strongly bends in back side, and producing a wide peri-vesical area around abdomen,,,,etc. Its characteristics are common to those of genus Zophoessa(generally included in the genus Lethe). It may be seen as a sharing of ancestral traits by both (various species of Zophoessa and Neope yama).
>Neope 属中最大的物种。 雄性生殖器的penis苍短,左右宽,背部强烈弯曲,在腹部周围形成宽阔的peri-vesical area,等。 其特征与Zophoessa属(通常包括在Lethe属中)的特征相同。 它可以被视为两者(Zophoessa 的各物种和 Neope yama)共享祖先特征。
附:日本産ヤマキマダラヒカゲNepoe niphonica(左)とサトキマダラヒカゲNeope goschkevitchii
写真(省略)ヤマキマダラヒカゲ屋久島亜種alt.1800m 2006.7.15/ サトキマダラヒカゲ東京都青梅市alt.200m 2021.5.14(春型)/同2021.8.22(夏型)alt.200m
種群Species group A(暫定temporary:niphonica-bremeri-agrestis)の撮影地photographing
地図(省略)●Neope agrestis雲南省西北部および四川省峨眉山/●Neope bremeri四川省青城山および浙江省杭州市/●Neope watanabei台湾/●Neope niphonica日本列島
種群Species group B(暫定temporary:goschkevitschii-pulaha complex-oberthueri)の撮影地
地図(省略)●Neope oberthueri雲南省西北部/●ユンナンクロキマダラヒカゲ雲南省南部/●Neope plahoidesベトナム北部/●未同定種(ramosa?)四川省成都西郊山地/●Neope ramosa浙江省天目山/●Neope didia(Neope pulaha台湾/●Neope goschkevitchii日本列島
前頁地図に示した撮影地点(丸ドット)や地図の範囲外におけるNeope属全体の分布は、北は北海道(南千島とサハリンの一部を含む)西はヒマラヤ地方、南は海南島やインドシナ半島北部に及ぶ。朝鮮半島など日本海の対岸地域、中国大陸の北部には分布しない。いわゆる「東亜半月孤」分布様式だが、屋久島を除く南西諸島には分布を欠く。Distribution of entire genus Neope, except for photographing points (circle dots) and outside range of previous map Hokkaido (including part of South Chishima and Saharin) in north,Himalayan region in west, and Hainan Island and northern part of Indochina in south. It is not distributed Korean and northern part of mainland China. also lacks distribution in South-west Islands except Yakushima. 除了拍摄点(圆点)和之前地图的范围之外 Neope 的分布,北是北海道(包括南千岛和沙哈林的一部分),西接喜马拉雅地区,南接海南岛和印度支那北部。不分布于朝鲜半岛等日本海对岸及中国大陆北部。 除屋久岛西南诸岛也缺乏分布。
やや特殊な数種を除く大陸産各種を、種群Aと種群Bに分けた。分類指標としてとりあえず重視したのは、雄交尾器の形状と下記の「特徴形質1」。ユンナンクロキマダラヒカゲは暫定的にBに含めたが、①の特徴に関してはAと相同。これらの組み合わせが必ずしも系統関係を反映しているわけではないが、Neope属の分布圏のほぼ全域で日本産2種同様に類似2種が混生していることは確かなようである。Except few species, divided into species group A and species group B. Focused male genitalia and "trait 1". 金平1995.4.8 included in B, but characteristics of ① homologous to A. Although these combinations do not necessarily reflect phylogenetic relation, it seems certain that 2 similar species coexist in entire distribution area of Neope. 除特殊物種,分为A群(包括日本的niphonica)和B群(包括日本的goschkevitschii)。专注于雄性生殖器和“特征 1”。金平1995.4.8是暂B,但①的特征与A同。些组合并不一定反映系统发育关系,但在Neope属的乎整个分布区域中两个相似的物种共存。
特徴形質①:種群A(左)と種群B(右)① Trait①:Species group A (left) and species group B (right)
写真(省略)後翅裏基部近くの3つの丸紋の並び方。左N.agrestis(buremeri種群の各種を通して共通)。最下方の紋が外側に大きく外れる。右N.oberthueri(pulaha種群の各種を通し傾向的に共通)=最下方一紋は特に目立って外側に外れない。3 round crests near the base of the back of the hind wings. Left N.agrestis (same as all bremeri species group) = bottom one is largely off to the outside. Right N.oberthueri (same as pulaha complex) = bottom one is not largely off to outside. 基部3圆。左: 田园荫眼蝶(bremeri 物种组全相同)= 底部的一个在外面。右:奥荫眼蝶(pulaha 复合种全相同)= 底部的一个不是在外面。
特徴形質②:左は「種群」に関わらず中国西南部高地帯の2種に共通。Trait②:Left is common to 2 species in Southwest China highlands regardless of any species group. 左是西南高地的两个物种所共有的,不分物种群。
写真(省略)(参考)後翅裏の亜外縁の眼状紋配列に関しては雲南省西北部に混棲する2種が共通し、上から2個目と3個目は緩やかに連なる:左N.agrestis(N.oberthueriも同様)。他の多くのNeope属の種では、上から3個目は2個目と顕著に外れる:右N.bremeri(agrestisとoberthueriを除く主要各種も同様)。(Reference) Arrangement of 3 eye-spots of near base of hind wing. Left: Neope agrestis. 2nd and third from top are loosely connected (same as Neope oberthueri that coexist in the northwestern part of Yunnan). Right: Neope bremeri (almost species of Neope except agrestis and oberthueri). Third one is outside.
(参考) 后翅近基部3个眼点的排列。左:agrestis、第二个和第三个是松散连接的(与云南西北部并存的oberthueri 相同)。右:bremeri(除agrestis 和oberthueri 之外全Neope)、第三个在外面。
梅里雪山に於けるNeope agrestisとNeope oberthuer
写真(省略)上左と中:Neope agrestis/上右端と下4頭:Neope oberthueri 雲南省梅里雪山明永(2300m)2009.6.11/左:Neope oberthueri右:Neope agrestis 雲南省梅里雪山明永(2300m)2009.6.11
写真(省略)この部分の紋の並び方に注目してください(説明は前頁にあります)。Please attention to arrangement of crests in this part (explanation is previous page). 请注意这部分的波峰排列(说明见前页)。
梅里雪山周辺のキマダラヒカゲ類:Ⅰ Neope agrestis 田园荫眼蝶
写真(省略24枚)
四川省峨眉山alt.2400m 1990.6. 1
雲南省白水台alt.2400m 2009.6.3
雲南省梅里雪山明永alt.2300m 2013.5.8
雲南省梅里雪山明永alt.2300m 2013.5.8
雲南省香格里拉~白水台alt.3200m 2005.6.19
雲南省香格里拉~白水台alt.3200m 2005.6.19
雲南省虎跳峡-香格里拉alt.2900m 2009.6.1
雲南梅里雪山明永alt.2300m 2009.6.5
雲南省梅里雪山明永alt.2300m 2009.6.5
雲南省梅里雪山明永alt.2300m 2012.6.29
雲南梅里雪山明永alt.2300m 2012.6.29
雲南省梅里雪山明永alt.2300m 2009.6.5
雲南省梅里雪山明永alt.2300m 2009.6.6
雲南梅里雪山明永alt.2300m 2012.6.29
雲南省梅里雪山明永alt.2300m 2017.6.15
(左と同一個体same as the left)*
雲南省梅里雪山明永alt.2300m 2017.6.14
雲南省梅里雪山明永alt.2300m 2017.6.14
雲南省梅里雪山明永alt.2300m 2017.6.15
雲南省梅里雪山明永alt.2300m 2017.6.15
雲南省梅里雪山明永alt.2300m 2017.6.15
雲南省梅里雪山西当‐雨崩(峠頂)alt.3900m 2009.6.11
雲南省維西東南方alt.2500m 2010.5.18
雲南省維西東南方alt.2500m 2010.5.18
*向きを統一するため反転した画像を含む/Includes flipped images to unify orientation/包括反转图像以统一方向
梅里雪山周辺ののキマダラヒカゲ類:Ⅱ Neope oberthueri 奥荫眼蝶
写真(省略16枚)
雲南省梅里雪山明永alt.2300m 2013.5.8
(左と同一個体same one as the left)*
雲南省梅里雪山明永alt.2300m 2013.5.10
(左と同一個体same one as the left)*
雲南省梅里雪山明永alt.2300m 2013.5.7
雲南省虎跳峡-香格里拉alt.2900m 2009.6.1
雲南省虎跳峡-香格里拉alt.2900m 2009.6.1
雲南省虎跳峡-香格里拉alt.2900m 2009.6.1
雲南省梅里雪山西当‐雨崩alt.3500m 2009.6.11
雲南省梅里雪山西当‐雨崩alt.3300m 2009.6.11*
雲南省梅里雪山明永alt.2300m 2017.6.15
(左と同一個体same one as the left)*
雲南省梅里雪山明永alt.2300m 2017.6.14
雲南省梅里雪山明永alt.2300m 2012.6.29
雲南省梅里雪山明永alt.2300m 2012.6.30
雲南省梅里雪山明永alt.2300m 2012.7.1
*向きを統一するため反転した画像を含む/Includes flipped images to unify orientation/包括反转图像以统一方向。
キマダラヒカゲ類三種の原記載図など Neope agrestis
写真(省略)
Satyrus Agrestis Oberthür= Neope agrestis (Oberthür, 1876)(a: upperside, b: underside)
Leech 1892-1893 Neope agrestis albicans
写真(省略)
梅里雪山明永(管理事務所内にて)2017.6.15
写真(省略)
Neope agrestis(左:雄2013.5.9梅里雪山/中:雌2009.6.1香格里拉‐虎飛峡/右:雄2012.6.29梅里雪山)
写真(省略3枚)
写真(省略8枚)
各地のNeope argestis-bremeri種群:1~6 Neope argestis/7&8 Neope bremeri 1雲南省梅里雪山雨崩 標高約3900m地点 2009.6.11/2雲南省梅里雪山明永 標高約2300m地点2017.6.15/3雲南省維西 標高約2500m地点/4雲南省香格里拉~虎跳峡 標高約2900m地点/5雲南省香格里拉~白水台 標高約3500m地点/6四川省峨眉山 標高約2400m地点/7四川省青城山 標高約900m地点/8浙江省杭州市街地(湖畔)標高約10m地点(female)
キマダラヒカゲ類三種の原記載図など Neope oberthueri & Neope similens
写真(省略)
Neope obertueri Leech 1891
Neope simulans Leech 1891
写真(省略)
Neope obertueri 雲南省梅里雪山 2009.6.11
Neope simulans 雲南省梅里雪山 2017.6.15
Neope obertueri雲南省香格里拉‐虎跳峡 2009.6.1
Neope obertueri(female?)雲南省梅里雪山 2013.5.10
Neope obertueri梅里雪山 2013.5.7
Neope obertueri梅里雪山 2013.5.10
Neope simulans梅里雪山 2017.6.15
翅表に関しては、oberthueri(変異の幅は広い)とsimulansの間に有意差は無いように思われる。Front wing no significant difference between oberthueri (have many variation) and simulans .翼桌 oberthueri(有很多变化)和 simulans 之间没有差异。
注1:開翅表示は全てサンプリング個体。All open wing displays are sampled individuals. 所有开放式机翼展示均为采样个体。
注2:上に紹介した大半の個体は雌雄の判別を行っていない。おそらく大多数は雄。確実に雌と分かる個体のみ、それを記した。
Most of individuals introduced above don’t distinguish between male and female. Probably majority are male. Only individuals that is surely identified as females are shown so. 上面介绍的大多数不分雌雄。可能大多数是雄性。确定为雌性的个体才会显示。
写真(省略)
2017.6.15。梅里雪山明永氷河下の入山管理署室内の床や窓際には、多くのキマダラヒカゲ類の死骸が散乱していた。agrestis, oberchueriの他、simulansも混じっていた。新鮮個体が多く、なぜ屋外に逃げ出さなかったのか不思議である。上:窓際にとまるoberthueri。下:床に転がっていたagrestisとsimulans(右から2頭目)。/Many dead Neope (including simulans) were scattered on floor and near windows of mountain climbing control office. Many are fresh, why they didn't escape outdoors? Above: oberthueri perching by window. Bottom: agrestis and simulans (2nd from right) lying on the floor./许多荫眼蝶的尸体散落在梅里雪山入山行政办公室的地板和窗户附近。很多都是新鲜的,为什么不逃到户外呢?上图:奥荫眼蝶栖息在窗边.。下图:田园荫眼蝶和(右二)拟网纹荫眼蝶躺在地板上。
写真(省略)
左left:Neope oberthurei(2009.6.2)/中center:Neope simulans/右right:Neope agrestis
写真(省略)
左left:Neope simulens拟网纹荫眼蝶/右right:Neope agrestis田园荫眼蝶和
写真(省略)
左left:simulans/中と右center and right:agrestis【↓左left:oberthueri/右right:agrestis】
写真(省略)
22017.6.15。全て梅里雪山管理事務所(上左を除く)。一部は別頁にも重複掲載。All photos(except top left)of this page are same data, and some photos are also introduced on another page.全是梅里雪山管理办公室(不含左上)。 有些在另一页上重复。
Neope agrestis/Neope oberthueri
写真(省略)
Neope agrestis 2012.7.9
Neope oberthueri 2013.5.10
氷河末端から数km下った流れの横にあるこの水溜まりに、100種近い蝶が水を吸いに訪れる(水面が黄緑色の藻で覆われ、多くの蝶が藻に捕まって溺れる)。指を差し出すと、様々な蝶が指にとまって吸汁する。ただしキマダラヒカゲ属2種は、この水溜まりよりも、周辺の路上で吸水していることのほうが多い。/Nearly 100 species butterflies come here to suck water in this puddle next to stream down 明永glacier terminus (surface of water is covered with yellow-green algae, many butterflies are drowned in here). Various butterflies are perch my finger and suck moisture. 2 species Neope also can see on the roads around puddle. /近100种蝴蝶来这里在明永冰川终点旁的水坑里吸水(水面长满黄绿藻,很多蝴蝶都淹死在这里)。各种各样的蝴蝶栖息在我的手指上吸湿。2种荫眼蝶也可以看在水坑周围的道路上。
Neope agrestis
写真(省略)
with Delias subnubila 雲南梅里雪山Alt.2300m 2012.7.9
写真(省略)
with Pieris dubernardi)2012.7.9 下右:雲南省維西東南方alt.2500m
Neope agrestis
写真(省略)
雲南省梅里雪山西当-雨崩(峠頂)alt.3900m 2009.6.11
これまで撮影した中での最高標高地。Neope as highest altitude taken picture.最高海拔拍摄的荫眼蝶的照片。
写真(省略)
雲南省維西東南方alt.2500m 2010.5.18
Neope agrestis
写真(省略)
雲南省維西東南方alt.2500m 2010.5.18
写真(省略)
雲南省梅里雪山雨崩alt.3500m付近、2009.6.11
Neope bremeri
写真(省略)
【上写真】四川省都江堰市青城山1989.4.16 。同じ場所には、雄前翅表の性標を欠き、後翅裏面3丸紋の3個目が特に大きく外れないpulaha系の種(ramosaまたはpulahoides?)も棲息。In the same place, live also pulaha-type species(ramosa or pulahoides?) that lacks sex patch on front wing surface, and 3 round crests near base of hind wing does not come off significantly. 在同一个地方,还生活着前翅表面没有性斑的pulaha型种(ramosa或者pulahoides?),后翅基部附近的3个圆嵴连接顺畅。
【下写真】浙江省杭州市内(西湖畔)1989.3.31