








The vicarious species of Neope niphonica and Neope goschevitschii (endemic species of Japan)found outside of Japan
大都市都心部を含めた日本の広い範囲(幾つかの離島を除く)に分布するサトキマダラヒカゲNeope goschevitschiiと、主に標高1000m以上の山岳地帯(寒冷地および房総半島では低標高地にも)に分布するヤマキマダラヒカゲNeope niphonicaは、ともに日本列島(サハリンの一部などを含む)の固有種である。
キマダラヒカゲ属は、朝鮮半島や中国北部、ロシア沿海地方など日本海対岸地方には分布せず、日本列島産2種のほか、台湾に4種、中国大陸(中部以南)・ヒマラヤ地方・インドシナ半島北部などに10種前後が分布することになっている。記載された種や亜種はかなりの数に上ると思われるが、相互の関係についての検証は全く成されていないというのが現状である。
筆者(青山は)以前、中国東部(杭州市)産のタイワンキマダラヒカゲ原名亜種Neope bremeri bremeriについて観察を行い、また「中国のチョウ」(東海大学出版会1998年)で、その時点で知り得たNeope属各種の雄外部生殖器の形状を比較し、各種間の相互関係を暫定的に考察した(166-168頁、315-322頁)。その後の15年間に、新たな知見の追加集積が成されているが、未だ総合的な纏めを行うには至っていない。最大の課題である日本産2種、ことにサトキマダラヒカゲの対応種についても、未解明のままである。
しかし、ひとつ、かなりの高い確率で指摘出来そうな事実がある。最終的な結論は将来の実証結果に委ねるとして、その概要をごく簡単に述べておく。
その前にまず、ヤマキマダラヒカゲ(以下“ヤマ”)とサトキマダラヒカゲ(以下“サト”)の外観上の区別点について説明しておこう。多くの形質による比較が成されているが、両種を確実に区別しうる安定的な指標形質は次の3点。
●①前翅外縁。
ヤマでは上部で緩く内側に湾曲、サトは湾曲しない。
●②前翅表第〇室眼状紋中の黒点。
ヤマで外縁寄りに、サトではほぼ中央部に位置する(♂についてのみ検証可)。
●③後翅裏基部の3個の丸紋の並び方。
ヤマでは下の一個が上2個の延長線上から明瞭に外側へ外れ、中央の一個からの距離は上の一個に比べて明らかに遠い。サトでは3個がほぼ均等の距離を保って並び、下の一個が外側へ大きく外れることはない。
上記3形質以外については傾向的な差異に過ぎず、それらに注目することで、しばしばかえって混乱してしまい、分からなくなってしまう。上記3形質は極めて安定的なので、慣れればそれによって100%区別が可能である。
それらのうち、①②に関しては、幾分印象による判断が必要とされるため、よほどの熟練者でなければ同定を誤ることがある。しかし③については特徴が安定的で、この形質のみで同定が可能となる(それに①②による検証を加味すれば、ほぼ間違いなく正しい同定が出来ると思う)。
*両種の幼生期の形態には著しい差異があり、また染色体数も異なることから、外観上の酷似とはうらはらに、思いのほか血縁の離れた関係にある可能性も考えられる。
中国大陸産各種のうち代表的な種について、大雑把に整理しておく(詳しくは「中国のチョウ」を参照されたし)。
日本産2種から最も類縁が遠いと思われるのが、オオキマダラヒカゲNeope yama。外部生殖器は構造的にZophoessa属との類似性も示され、Neope属の中ではかなり離れて位置付けされる可能性がある。
Neope属中、低標高地を含む最も広い範囲に分布するウラキマダラヒカゲNeope muilheadiiも、外観が他の多くの種と顕著に異なる。♂外部生殖器にも、uncusの先端が2分岐するという他の各種に見られない明確な特徴を示す。ただし基本構造的には他各種と共通し、外観の相違から判断されるほど特異な存在ではないのかも知れない(将来複数種に分割される可能性もある)。分布状況や生態的地位に於いては、この種がサトキマダラヒカゲの代置的存在にあるように思われるが、形態形質的な面での相関性は特に見出し得ない。
シロキマダラヒカゲNeope armandiは、山岳地帯を中心として、やはり広い範囲に分布している。ウラキマダラヒカゲに於いても同様のことが言えるが、季節・雌雄・地域などに於ける外観上の差が著しく、後翅表に白色やオレンジ色部分が広がり他の各種とは顕著に異なって見える個体から、意外に一般のキマダラヒカゲに類似した個体まで、多岐に亘っている。♂外部生殖器にも、他の種から隔てられる顕著な特徴は見出されず(相対的にvalva基半部の幅が狭いというのが特徴と言えば特徴と言える)、外観で想像される以上に、以下に述べる各種に、かなり近縁な存在であるのかも知れない(やはり複数種から成る可能性あり)。
ヤマやサトの対応種は、(Neope simlansなど幾つかの特異な種を除く)残る各種の中に見出されるものと思われる。上記の①②の分類指標形質を見る限りに於いては、それらの全ての種がヤマに対応するように思われ、サトに対応すると積極的に評価しうるような集団は見出されない。四川省から記載されたNeope shirozuiをサトの対応種と考える意見もあるが、おそらくはN.bremeriやN.agurestis(あるいはN.pulaha)に収斂される、顕著な変異型(または地域集団)ではないかと考えている。
キマダラヒカゲ属の分類に当って最も問題となる事項のひとつは、♂外部生殖器の形状比較による考察が困難なことである。基本的な構造に於いて種間の差が少なく、かつ種内に於ける末端的な部分での変異が多様なことから、外部生殖器の形状から種間関係の正確な実態を把握するのは至難の業なのである。
ヤマとサトの外部生殖器の形状にも、ほとんど差異がない。そのため両者は、(上記したような)血縁上相当に離れた関係にあるとする見解とは逆に、比較的新しい時代に分化した集団ではないか、という見解も成し得よう。
ヤマとサトの間のみでなく、(yama, muilheadiiなど明らかな固有の特徴を示す種を除けば)国外産の各種間にも、基本的な構造差は、ほとんどないと言ってよい。差異が見られるのは下記のような部位。
●①プロポーション(uncusやvalvaやpenisの相対長)。
●②末端形質(valvaの先端部分など)。
●③penisの鋸状突起。
このうち、①や②については、種(または地域集団)間である程度の安定差が示されるが、③については、同一種の個体間(しばしば同一地域集団の個体間)でも、多様な表現が成される。また①については、しばしば亜種間(ことにN.bremeri watanabeiとN.bremeri bremeri間)で顕著な差が示される。
各部位が大型で、valvaやuncusの前後長が長く、penisの鋸歯群が最も発達するのが、タイワンキマダラヒカゲ台湾亜種N.bremeri watanabei。逆に各プロポーションが最もコンパクト(前後に寸詰まり)で、penisの鋸歯も発達が弱い(あるいは消失する)のがクロキマダラヒカゲN.agrestis。タイワンキマダラヒカゲ原名亜種(アカキマダラヒカゲ)N.bremeri bremeriと日本産の2種は、ほぼ中間的な段階を示し、アリサンキマダラヒカゲN.pulaha didia(および原名亜種を含む大陸産の幾つかの近似分類群)も、ほぼそれに準じる(penisの鋸歯の発達はおおむね弱い)。
思うに、外部生殖器の形状(例えば全体のプロポーションやpenisの棘の出現程度を含む末端部分)の差異は、種間差というよりも、種内(地域集団間、ときによっては個体間)変異と見るのが妥当ではないかと。
外観(翅の概形・斑紋・色彩など)の差異についても同様のことが言えるであろう。外観や外部生殖器の形状が酷似する複数の(しかし少数の)“種(または上種)”が存在、それぞれの種内で(同一空間に於いて並行的に相互相似する)変異の幅を示すことから、多くの種から成るように見えるのである(♂の性標の表現の違いにどのような意味があるのか? 今後の重要な検証課題だが、あえて誤解を顧みずに言えば、上記した他の形質同様に、2次的かつ並行的な相似/相違である可能性も考えられる)。
いずれにしろ、外部生殖器の形状に斑紋など外観的特徴を加味して考察すれば、国外産の典型Neope各種は、全ての種がヤマに繋がるように思え、サトに繋がる種は存在しない、と言い切ってもよさそうなのだが、今一度、外観上に表現された形質に惑わされることなく、改めて整理検討しておきたい。
上記したように、私見では、典型Neopeはごく少数の種、ほぼ同一の(かつ地域集団などの別に於いて様々な段階で異なった表現が成される)形質を備えた2つの分類群(種または上種)から成っているのでは、と見ている。ヤマもサトも、各々そのどちらかに収斂されるのである(同一空間に存在することによる偶然または必然的な相似)。
そのことを示唆する例を挙げておく。どうやらNeope分布圏内に於いては、幾つかの地域で(yama, muilheadii, armandiなどを除き)日本のヤマ&サト同様に、2つの分類群(種または上種)が混在している可能性がある。「2つの」ということが重要で、決して「多数の」ということではない。日本のヤマ同様に、いや遥かにそれを上回る顕著な変異を示すことから、「2つ」ではなく「多数」に見えるのだが、それらの「多数」は、全て「2つ」に収斂されるわけである。
その2つとは、すなわち「タイワンキマダラヒカゲNeope bremeri」と「アリサンキマダラヒカゲNeope pulaha」。ヤマが前者に対応することは容易に理解出来ようが、思い切って言うならば、サトは後者に対応する(後述)。
前者には、中国中部~東部のN.bremeri原名亜種や、西南部山岳地帯産のN.agrestisが含まれる。以下にこれまでに撮影した個体を、雲南省産N.agrestisを中心に紹介しておく。

Neope agrestis 2013.5.8 雲南梅里雪山Alt.2300m

Neope agrestis 2013.5.8 雲南梅里雪山Alt.2300m

Neope agrestis 2010.5.18 雲南維西Alt.2200m

Neope agrestis 2010.5.18 雲南維西Alt.2200m

Neope agrestis 2010.5.18 雲南維西Alt.2200m

Neope agrestis 2009.6.5 雲南梅里雪山Alt.2300m

Neope agrestis 2009.6.5 雲南梅里雪山Alt.2300m

Neope agrestis 2005.6.19 雲南香格里拉~白水台Alt.3200m

Neope agrestis 2005.6.19 雲南香格里拉~白水台Alt.3200m

Neope agrestis 2005.6.11 雲南梅里雪山Alt.3500m

Neope agrestis 2005.6.12 雲南梅里雪山Alt.3600m

Neope agrestis 2012.6.29 雲南梅里雪山Alt.2300m

Neope agrestis 2012.6.29 雲南梅里雪山Alt.2300m

Neope agrestis 1990.6. 1 四川峨眉山Alt.2400m

Neope sp. 1995.4. 14 雲南金平県Alt.2000m

Neope agrestis 2012.7.9 雲南梅里雪山Alt.3500m

Neope agrestis 2009.6.3 雲南白水台Alt.2400m

Neope agrestis 2009.6.5 雲南梅里雪山Alt.2300m

Neope agrestis 2005.6.12 雲南梅里雪山Alt.3600m
四川峨眉山産と雲南金平産を除き、いずれも雲南北部の高標高地産で、5月上旬から7月上旬に至る撮影個体。8月中旬から10月上旬にかけての探訪時には確認していず、この期間に発生しているものと思われる。同一地点(例えば梅里雪山明永氷河末端下部の標高2300m地点)に於いても丸2ヶ月の期間に亘るため、2世代を繰り返している可能性も考えられなくはないが、5月上旬の個体と6月下旬の個体間に有意な差が見られないことから、年1化の発生(長期間に亘って暫時出現)と考えたほうが妥当であろう。もし2世代を繰り返しているとすれば、世代間の形質差がほとんどないということになる。(しばしばヤマキマダラヒカゲ房総亜種N.niphonicakiyosumiensis春型個体に見られるように)翅型がコンパクトで寸詰まりな印象、外縁の白色鱗と翅脈末端の黒色鱗のコントラストが強いため、外縁が顕著に凹凸を帯びて見える傾向がある。
全体的な形質はタイワンキマダラダラヒカゲ原名亜種(アカキマダラヒカゲ)N.bremeri bremeri春型に連続し、日本のヤマキマダラヒカゲN.niphonicaとも繋がるように思われる。写真下2枚の個体のように、裏面地色の色調がぼやけて明るい赤茶色を帯びた個体なども見出されるが、個体変異の範疇に入るものと考えている。「中国のチョウ」で“ウンナンクロキマダラヒカゲ”として紹介した雲南省南部のベトナム国境に近い金平産の個体は、北部のagrestisとは大分印象がことなり、bremeriに所属せしめるべきかも知れない。N.pulahaの系統とも考えられるが、裏面基部の丸紋の並び方は、後述するpulahaと仮同定した雲南北部山岳地帯産と異なり、明らかにagrestis~bremeri~niphonicaと共通する特徴を示している。

Neope bremeri bremeri春型♂ 1989.3.31浙江杭州市Alt.100m

Neope bremeri bremeri春型♀ 1989.3.31浙江杭州市Alt.100m

Neope bremeri bremeri春型♂ 1989.4 .15 四川青城山Alt.800m

Neope bremeribremeri夏型♂ 1989.8. 4 四川青城山Alt.800m

Neope pulahoides?♂ 1990.4. 8 四川青城山Alt.800m

Neope bremeri bremeri? 夏型♀ 1990.6. 1 四川峨眉山Alt.2400m
♂外部生殖器のプロポーションや末端構造に於いて(あるいは翅の概形や斑紋・色彩などに於いても)、クロキマダラヒカゲNeope agrestisと対極的な印象を持つのが、タイワンキマダラヒカゲ台湾亜種(N.bremeri watanabei)である。N.bremeriの大陸産原名亜種と台湾亜種の間には、一般的常識による種間差を優に越える差があるように見受けられるが、形質の一定方向への“傾き”であると捉えれば、両者を同一種として扱うことは妥当であると思われる。ただしその場合、反対方向の傾斜上に位置するN.agrestisの処遇も考慮しなければならない。大陸東部(杭州)産N.bremeri原名亜種は、全体の傾斜の中で、台湾亜種N.bremeri watanabeiと、大陸西南部高地産 N.agrestisとの中間に位置する、と言ってよい(日本産のN.niphonicaについてもそれに近いことが言えそうである)。
四川成都市近郊(青城山)産もN.bremeriに含めた。杭州市産と同じくbremeriの原名亜種に置く事が妥当か否かについては今後の詳しい検証を待たねばならないが、盛夏に同一種の夏型(外部生殖器形状の一致を検証済み)が出現することから、夏型の出現しないN.agrestisとは一線を区し、(青城山に隣接した西嶺雪山で撮影した夏型♀共々)狭義のbremeriに含めておく。
ほかに青城山からは、性票(特殊鱗の集中散布)を欠く春型♂を記録していて、「中国のチョウ」では暫定的にN.pulahoidesとしておいた。N.pulahoidesやN.pulahinaは、広義にはN.pulahaの一群に所属するものと考えられるが、この個体は、あるいはpulaha ではなくbremeriに繋がるのかも知れない。先に記したように、性標(特殊鱗の有無や散布範囲)が種間関係に於いてどのような意味を持ち、分類指標としてどの程度の重要性を示すのかを検証することは、今後の重要な検証課題であると思われる。
それらの課題はさておき、(N.niphonicaの分類上の処遇はともかく)N.bremeri, N.agrestis, N.niphonica等の各種が、同一の形質傾斜上に置かれた、ひとつの上種super-speciesを形成する可能性を、一応の結論としておきたい。
では、サトキマダラヒカゲNeope goschevitschiiは、どの傾斜上に位置するのであろうか? 筆者の結論は、少々乱暴ではあるが「アリサンキマダラヒカゲN.pulaha」が、国外に於ける日本固有種「サトキマダラヒカゲ」の対応種であるということ。
“乱暴”の実態を補足しつつ、中国西南部に於ける「サトキマダラヒカゲ」対応集団の写真を紹介していく。

Neope pulaha【仮同定】(手前)とN.agrestis(奥)2012.6. 29 雲南梅里雪山Alt.2300m

Neope pulaha【仮同定】 2013.5. 7 雲南梅里雪山Alt.2300m

Neope pulaha【仮同定】 2013.5. 8 雲南梅里雪山Alt.2300m

Neope pulaha【仮同定】 2013.5. 8 雲南梅里雪山Alt.2300m

Neope pulaha【仮同定】 2013.5. 8 雲南梅里雪山Alt.2300m(上と同一個体)

Neope pulaha【仮同定】 2013.5. 10 雲南梅里雪山Alt.2300m

Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m

Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m

Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m

Neope pulaha【仮同定】 2009.6. 11雲南梅里雪山Alt.3500m

Neope pulaha【仮同定】 2009.6. 11雲南梅里雪山Alt.3500m

Neope pulaha【仮同定】 2012.6. 30雲南梅里雪山Alt.2300m

Neope pulaha【仮同定】 2012.6. 29雲南梅里雪山Alt.2300m(12枚前と同一カット)
現時点では(比較的限られた地域で広範囲とはいえないが)少なくとも4つの地域に於いて“典型Neope”と呼ぶべき2種の混棲が知られている。先にも述べたように、重要なことは、単に2種が混棲していると言うことよりも、それらの地域に見られる(一見多様な変異形質を示す)“典型Neope”の全てが、そのどちらかの種に帰属する、ということである。
ひとつはもちろん日本産の2種。そして台湾のタイワンキマダラヒカゲ台湾亜種Neope bremeri watanabeiとアリサンキマダラヒカゲ台湾亜種Neope pulaha didia。以下に述べる雲南省北部山地に於けるクロキマダラヒカゲNeope agrestisとNeope pulaha類似種。およびそれと同一の組み合わせと考え得るミャンマー北部産の2種(Butterflies Musiam展示の過程で多数の標本を検証)。それ以外の地域では、今のところ前者(bremeriなどniponica対応集団)のみしか確認していないが、より多くの地域で“2種”が混棲している可能性も有り得る。
先に検証したように、niphonica~bremeri~agrestisは、同じ傾斜上に位置するniphonica対応集団と考えられる。ならば、一方のgoschchevitschii~pulaha didia(台湾亜種)~大陸東南部産pulaha類似種も、同じ傾斜上に置かれる対応集団、と考えるのは早計に過ぎるであろうか?
常識的には、暴論には違いない。アリサンキマダラヒカゲ台湾亜種N.pulaha didiaは(中国大陸産アリサンキマダラヒカゲN.pulahaのいくつかの亜種ともども)、外観上、様々な点でサトキマダラヒカゲN.goschevitschiiとは対極的な特徴を示している。 ヤマとサトの有意な区別点たる、「前翅外縁のカーブの仕方」「前翅表第0室眼状紋内の黒点の位置」「後翅裏面基部の3個の丸紋の並び方」など、いずれもサトとの共通性は全く示さず、むしろずっとヤマ的だ。アリサンとサトは、“典型Neope”の中では最も離れた位置にあると考えるのが妥当であろう。
大陸西南部高地産のN.pulaha類似種に於いても、以下に述べる斑紋などの幾つかの特徴はともかく、より広域に分布するN.bremeri~N.agrestisに対して、明らかに(高標高地のみに)分布域が限られている。日本のヤマとサトの関係とは正反対なのである。
しかし、日本と中国では、近縁種間の組み合わせに於いて生態的地位が逆転する、という現象は、枚挙にいとまない。また、表現された形質が、必ずしも系統を反映しているとは限らない、ということも、多くの例で知られるところである。
そして、非常に気になる事実がひとつ。最初に示した、ヤマとサトの、最も安定的な分類指標である「③後翅裏面基部の3つの丸紋の並び方」に於いて、大陸西南部(雲南省北部)山地産のN.pulaha類似種と、日本産N.goschevitschiiは、見事に一致するのである。
そのような前提で改めてこのN.pulaha類似種に注目すれば、裏面の色調や全体的な印象も、どこかサトに共通する印象が感じられる(ただし現地での観察に於いて、ことに飛翔時には本種は真っ黒に見え、一方、筆者が「中国のチョウ」で“クロキマダラヒカゲ”と名付けたNeope agrestisは、遥かに明るい色調~赤褐色~に見えることから、和名については再検討するべきと考えている)。
ちなみに、最初に記したごとく、♂外部生殖器の形状は、ヤマとサトの間で酷似し、全く相同のようにも見えるが、川副昭人氏も指摘し(若林守男氏との共著による「日本産蝶類大図鑑」保育社) 筆者も「中国のチョウ」で解説したように、非常にデリケートではあるが、それなりの安定差が見出される。すなわち、ヤマではpenisに生じる鋸歯群の個々の鋸歯が相対的に大型で、やや疎らに散布される傾向があるのに対し、サトでは個々の鋸歯がより小型で、大きさや散布密度が一定しているということ。また、(あくまで傾向的・印象的な相違に過ぎぬが)uncus概形の全体的な印象が、サトでは茫洋とした感じで、ヤマのような引き締まりに欠ける気がする。それらの傾向は、buremeri watanabei~bremeri buremeri~agrestisと、pulaha complexの間にも当て嵌まるように思える。
筆者は、原則、Neope分布圏内の多くの地域で、2組の“典型Neope”の組み合わせが成されているのではないかと考えている。そして、どの組み合わせの2つの種も、ヤマ、サト、それぞれの傾斜上に位置しているのではないかと。日本列島に於ける2つの種の酷似は、並行的に成された、2次的な類似だと解釈したい。
むろん、それぞれの組み合わせに於ける2種が、同じ傾斜上の同一種群間に帰属するのではなく、各地での組み合わせごとに、(少なくとも“サト”に対応する側は)別系統の集団からなっている可能性も否定出来ない。ここに示した雲南省北部山地産“アリサンキマダラヒカゲNeope pulaha類似種”が、果たして系統上、(台湾産亜種をはじめとした)ほかのアリサンキマダラヒカゲ各亜種に繋がるのかなど、全く分かっていない。
それらを含めた、より詳しい検証が成されなければならない。幼生期、生態、行動、生理の比較、性標(特殊鱗粉)の散布状況と機質的機能的意味の解明、むろんDNA解析による系統的考察。
頼りとするのが高橋真弓氏であることは、今も昔と変わらない。氏は今も(国内外に調査旅行に赴かれるなど)積極的に活動を続けていられるが、来年には古希(80歳)を迎えられる。高橋氏に全てを頼るというのは酷であろう。といって、小生(青山)や日本に於けるLethe族研究の第一人者である鈴木英文氏も、前期高齢者(65歳)に突入する。我々が成すことの出来る範囲は限られていよう。若い方々(中国をはじめとする国外の人々を含む)によるアグレッシブな探求を、切に願うところである。

Neope agrestis 2012.6. 29雲南梅里雪山Alt.2300m

Neope agrestis 2009.6. 1雲南香格里拉-虎跳峡Alt.2800m

Neope agrestis 2009.6. 1雲南香格里拉-虎跳峡Alt.2800m

Neope agrestis 2013.5. 9雲南梅里雪山Alt.2300m

Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m

Neope pulaha【仮同定】 2009.6. 11 雲南梅里雪山Alt.3500m

Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m

Neope pulaha【仮同定】 2013.5. 10 雲南梅里雪山Alt.2300m

Neope pulaha【仮同定】(下4頭と上右)/Neope agrestis(上左2頭) 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m

Neope pulaha【仮同定】(左)/Neope agrestis(右) 2013.5. 10 雲南梅里雪山Alt.2300m


Neope pulaha【仮同定】(左)/Neope agrestis(右)
追記
小生はコレクションの趣味を全く持たず、原則としていかなる地域でも採集は行わないが、Neopeをはじめ、Ypthima(ウラナミジャノメ属)、 Pieris(モンシロチョウ属)、 Parnara(イチモンジセセリ属)等々、ごく地味ではあるが、日本やアジアの蝶相(ひいては各地域のアイデンティティイ)の解明に当たって極めて重要な意味をもつ幾つかの分類群については、必要に応じて採集を行っている。
注1:大抵の場合、補虫網を使わず手で摘まんだり叩き落したり足で踏んづけたりして採取するので、コレクターの視点からの標本的価値は全くない。
注2:それらの標本は中国各地の博物館や来年度にオープン予定の「静岡県立博物館」などに保管。
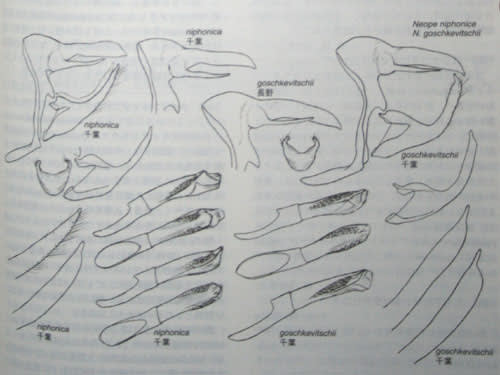


上左:goschevvitschiiとniphonica
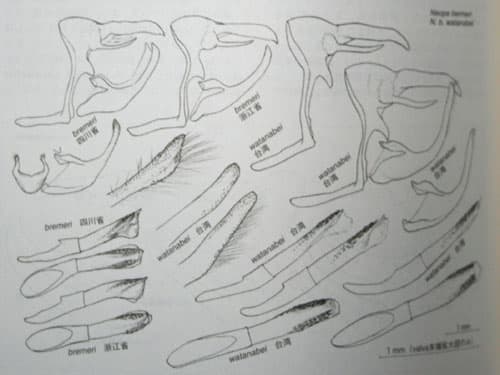
上右:bremeri bremeriとbremeri watanabei
下左:agrestis
下右:pulahaほか
【「中国のチョウ」より】
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
僕のブログのうち、いわば“趣味”でアップしている「ElvisとBeatlesの狭間で~Johnny Tillotsonの時代」や「朝と夜の狭間で~My Sentimental Journey」と異なり、昆虫や植物関係の話題については、“仕事”の一環と考えています。「青山潤三ネイチャークラブ」の会員諸氏や、それに準じる協力者諸氏に向けての発信です。(たまたま訪問された方はともかく)継続してブログを訪問される方には、各自何らかの形での自主的な援助を望んでいる次第です。その旨、ご承知おき下さい。
大都市都心部を含めた日本の広い範囲(幾つかの離島を除く)に分布するサトキマダラヒカゲNeope goschevitschiiと、主に標高1000m以上の山岳地帯(寒冷地および房総半島では低標高地にも)に分布するヤマキマダラヒカゲNeope niphonicaは、ともに日本列島(サハリンの一部などを含む)の固有種である。
キマダラヒカゲ属は、朝鮮半島や中国北部、ロシア沿海地方など日本海対岸地方には分布せず、日本列島産2種のほか、台湾に4種、中国大陸(中部以南)・ヒマラヤ地方・インドシナ半島北部などに10種前後が分布することになっている。記載された種や亜種はかなりの数に上ると思われるが、相互の関係についての検証は全く成されていないというのが現状である。
筆者(青山は)以前、中国東部(杭州市)産のタイワンキマダラヒカゲ原名亜種Neope bremeri bremeriについて観察を行い、また「中国のチョウ」(東海大学出版会1998年)で、その時点で知り得たNeope属各種の雄外部生殖器の形状を比較し、各種間の相互関係を暫定的に考察した(166-168頁、315-322頁)。その後の15年間に、新たな知見の追加集積が成されているが、未だ総合的な纏めを行うには至っていない。最大の課題である日本産2種、ことにサトキマダラヒカゲの対応種についても、未解明のままである。
しかし、ひとつ、かなりの高い確率で指摘出来そうな事実がある。最終的な結論は将来の実証結果に委ねるとして、その概要をごく簡単に述べておく。
その前にまず、ヤマキマダラヒカゲ(以下“ヤマ”)とサトキマダラヒカゲ(以下“サト”)の外観上の区別点について説明しておこう。多くの形質による比較が成されているが、両種を確実に区別しうる安定的な指標形質は次の3点。
●①前翅外縁。
ヤマでは上部で緩く内側に湾曲、サトは湾曲しない。
●②前翅表第〇室眼状紋中の黒点。
ヤマで外縁寄りに、サトではほぼ中央部に位置する(♂についてのみ検証可)。
●③後翅裏基部の3個の丸紋の並び方。
ヤマでは下の一個が上2個の延長線上から明瞭に外側へ外れ、中央の一個からの距離は上の一個に比べて明らかに遠い。サトでは3個がほぼ均等の距離を保って並び、下の一個が外側へ大きく外れることはない。
上記3形質以外については傾向的な差異に過ぎず、それらに注目することで、しばしばかえって混乱してしまい、分からなくなってしまう。上記3形質は極めて安定的なので、慣れればそれによって100%区別が可能である。
それらのうち、①②に関しては、幾分印象による判断が必要とされるため、よほどの熟練者でなければ同定を誤ることがある。しかし③については特徴が安定的で、この形質のみで同定が可能となる(それに①②による検証を加味すれば、ほぼ間違いなく正しい同定が出来ると思う)。
*両種の幼生期の形態には著しい差異があり、また染色体数も異なることから、外観上の酷似とはうらはらに、思いのほか血縁の離れた関係にある可能性も考えられる。
中国大陸産各種のうち代表的な種について、大雑把に整理しておく(詳しくは「中国のチョウ」を参照されたし)。
日本産2種から最も類縁が遠いと思われるのが、オオキマダラヒカゲNeope yama。外部生殖器は構造的にZophoessa属との類似性も示され、Neope属の中ではかなり離れて位置付けされる可能性がある。
Neope属中、低標高地を含む最も広い範囲に分布するウラキマダラヒカゲNeope muilheadiiも、外観が他の多くの種と顕著に異なる。♂外部生殖器にも、uncusの先端が2分岐するという他の各種に見られない明確な特徴を示す。ただし基本構造的には他各種と共通し、外観の相違から判断されるほど特異な存在ではないのかも知れない(将来複数種に分割される可能性もある)。分布状況や生態的地位に於いては、この種がサトキマダラヒカゲの代置的存在にあるように思われるが、形態形質的な面での相関性は特に見出し得ない。
シロキマダラヒカゲNeope armandiは、山岳地帯を中心として、やはり広い範囲に分布している。ウラキマダラヒカゲに於いても同様のことが言えるが、季節・雌雄・地域などに於ける外観上の差が著しく、後翅表に白色やオレンジ色部分が広がり他の各種とは顕著に異なって見える個体から、意外に一般のキマダラヒカゲに類似した個体まで、多岐に亘っている。♂外部生殖器にも、他の種から隔てられる顕著な特徴は見出されず(相対的にvalva基半部の幅が狭いというのが特徴と言えば特徴と言える)、外観で想像される以上に、以下に述べる各種に、かなり近縁な存在であるのかも知れない(やはり複数種から成る可能性あり)。
ヤマやサトの対応種は、(Neope simlansなど幾つかの特異な種を除く)残る各種の中に見出されるものと思われる。上記の①②の分類指標形質を見る限りに於いては、それらの全ての種がヤマに対応するように思われ、サトに対応すると積極的に評価しうるような集団は見出されない。四川省から記載されたNeope shirozuiをサトの対応種と考える意見もあるが、おそらくはN.bremeriやN.agurestis(あるいはN.pulaha)に収斂される、顕著な変異型(または地域集団)ではないかと考えている。
キマダラヒカゲ属の分類に当って最も問題となる事項のひとつは、♂外部生殖器の形状比較による考察が困難なことである。基本的な構造に於いて種間の差が少なく、かつ種内に於ける末端的な部分での変異が多様なことから、外部生殖器の形状から種間関係の正確な実態を把握するのは至難の業なのである。
ヤマとサトの外部生殖器の形状にも、ほとんど差異がない。そのため両者は、(上記したような)血縁上相当に離れた関係にあるとする見解とは逆に、比較的新しい時代に分化した集団ではないか、という見解も成し得よう。
ヤマとサトの間のみでなく、(yama, muilheadiiなど明らかな固有の特徴を示す種を除けば)国外産の各種間にも、基本的な構造差は、ほとんどないと言ってよい。差異が見られるのは下記のような部位。
●①プロポーション(uncusやvalvaやpenisの相対長)。
●②末端形質(valvaの先端部分など)。
●③penisの鋸状突起。
このうち、①や②については、種(または地域集団)間である程度の安定差が示されるが、③については、同一種の個体間(しばしば同一地域集団の個体間)でも、多様な表現が成される。また①については、しばしば亜種間(ことにN.bremeri watanabeiとN.bremeri bremeri間)で顕著な差が示される。
各部位が大型で、valvaやuncusの前後長が長く、penisの鋸歯群が最も発達するのが、タイワンキマダラヒカゲ台湾亜種N.bremeri watanabei。逆に各プロポーションが最もコンパクト(前後に寸詰まり)で、penisの鋸歯も発達が弱い(あるいは消失する)のがクロキマダラヒカゲN.agrestis。タイワンキマダラヒカゲ原名亜種(アカキマダラヒカゲ)N.bremeri bremeriと日本産の2種は、ほぼ中間的な段階を示し、アリサンキマダラヒカゲN.pulaha didia(および原名亜種を含む大陸産の幾つかの近似分類群)も、ほぼそれに準じる(penisの鋸歯の発達はおおむね弱い)。
思うに、外部生殖器の形状(例えば全体のプロポーションやpenisの棘の出現程度を含む末端部分)の差異は、種間差というよりも、種内(地域集団間、ときによっては個体間)変異と見るのが妥当ではないかと。
外観(翅の概形・斑紋・色彩など)の差異についても同様のことが言えるであろう。外観や外部生殖器の形状が酷似する複数の(しかし少数の)“種(または上種)”が存在、それぞれの種内で(同一空間に於いて並行的に相互相似する)変異の幅を示すことから、多くの種から成るように見えるのである(♂の性標の表現の違いにどのような意味があるのか? 今後の重要な検証課題だが、あえて誤解を顧みずに言えば、上記した他の形質同様に、2次的かつ並行的な相似/相違である可能性も考えられる)。
いずれにしろ、外部生殖器の形状に斑紋など外観的特徴を加味して考察すれば、国外産の典型Neope各種は、全ての種がヤマに繋がるように思え、サトに繋がる種は存在しない、と言い切ってもよさそうなのだが、今一度、外観上に表現された形質に惑わされることなく、改めて整理検討しておきたい。
上記したように、私見では、典型Neopeはごく少数の種、ほぼ同一の(かつ地域集団などの別に於いて様々な段階で異なった表現が成される)形質を備えた2つの分類群(種または上種)から成っているのでは、と見ている。ヤマもサトも、各々そのどちらかに収斂されるのである(同一空間に存在することによる偶然または必然的な相似)。
そのことを示唆する例を挙げておく。どうやらNeope分布圏内に於いては、幾つかの地域で(yama, muilheadii, armandiなどを除き)日本のヤマ&サト同様に、2つの分類群(種または上種)が混在している可能性がある。「2つの」ということが重要で、決して「多数の」ということではない。日本のヤマ同様に、いや遥かにそれを上回る顕著な変異を示すことから、「2つ」ではなく「多数」に見えるのだが、それらの「多数」は、全て「2つ」に収斂されるわけである。
その2つとは、すなわち「タイワンキマダラヒカゲNeope bremeri」と「アリサンキマダラヒカゲNeope pulaha」。ヤマが前者に対応することは容易に理解出来ようが、思い切って言うならば、サトは後者に対応する(後述)。
前者には、中国中部~東部のN.bremeri原名亜種や、西南部山岳地帯産のN.agrestisが含まれる。以下にこれまでに撮影した個体を、雲南省産N.agrestisを中心に紹介しておく。
Neope agrestis 2013.5.8 雲南梅里雪山Alt.2300m
Neope agrestis 2013.5.8 雲南梅里雪山Alt.2300m
Neope agrestis 2010.5.18 雲南維西Alt.2200m
Neope agrestis 2010.5.18 雲南維西Alt.2200m
Neope agrestis 2010.5.18 雲南維西Alt.2200m
Neope agrestis 2009.6.5 雲南梅里雪山Alt.2300m
Neope agrestis 2009.6.5 雲南梅里雪山Alt.2300m
Neope agrestis 2005.6.19 雲南香格里拉~白水台Alt.3200m
Neope agrestis 2005.6.19 雲南香格里拉~白水台Alt.3200m
Neope agrestis 2005.6.11 雲南梅里雪山Alt.3500m
Neope agrestis 2005.6.12 雲南梅里雪山Alt.3600m
Neope agrestis 2012.6.29 雲南梅里雪山Alt.2300m
Neope agrestis 2012.6.29 雲南梅里雪山Alt.2300m
Neope agrestis 1990.6. 1 四川峨眉山Alt.2400m
Neope sp. 1995.4. 14 雲南金平県Alt.2000m
Neope agrestis 2012.7.9 雲南梅里雪山Alt.3500m
Neope agrestis 2009.6.3 雲南白水台Alt.2400m
Neope agrestis 2009.6.5 雲南梅里雪山Alt.2300m
Neope agrestis 2005.6.12 雲南梅里雪山Alt.3600m
四川峨眉山産と雲南金平産を除き、いずれも雲南北部の高標高地産で、5月上旬から7月上旬に至る撮影個体。8月中旬から10月上旬にかけての探訪時には確認していず、この期間に発生しているものと思われる。同一地点(例えば梅里雪山明永氷河末端下部の標高2300m地点)に於いても丸2ヶ月の期間に亘るため、2世代を繰り返している可能性も考えられなくはないが、5月上旬の個体と6月下旬の個体間に有意な差が見られないことから、年1化の発生(長期間に亘って暫時出現)と考えたほうが妥当であろう。もし2世代を繰り返しているとすれば、世代間の形質差がほとんどないということになる。(しばしばヤマキマダラヒカゲ房総亜種N.niphonicakiyosumiensis春型個体に見られるように)翅型がコンパクトで寸詰まりな印象、外縁の白色鱗と翅脈末端の黒色鱗のコントラストが強いため、外縁が顕著に凹凸を帯びて見える傾向がある。
全体的な形質はタイワンキマダラダラヒカゲ原名亜種(アカキマダラヒカゲ)N.bremeri bremeri春型に連続し、日本のヤマキマダラヒカゲN.niphonicaとも繋がるように思われる。写真下2枚の個体のように、裏面地色の色調がぼやけて明るい赤茶色を帯びた個体なども見出されるが、個体変異の範疇に入るものと考えている。「中国のチョウ」で“ウンナンクロキマダラヒカゲ”として紹介した雲南省南部のベトナム国境に近い金平産の個体は、北部のagrestisとは大分印象がことなり、bremeriに所属せしめるべきかも知れない。N.pulahaの系統とも考えられるが、裏面基部の丸紋の並び方は、後述するpulahaと仮同定した雲南北部山岳地帯産と異なり、明らかにagrestis~bremeri~niphonicaと共通する特徴を示している。
Neope bremeri bremeri春型♂ 1989.3.31浙江杭州市Alt.100m
Neope bremeri bremeri春型♀ 1989.3.31浙江杭州市Alt.100m
Neope bremeri bremeri春型♂ 1989.4 .15 四川青城山Alt.800m
Neope bremeribremeri夏型♂ 1989.8. 4 四川青城山Alt.800m
Neope pulahoides?♂ 1990.4. 8 四川青城山Alt.800m
Neope bremeri bremeri? 夏型♀ 1990.6. 1 四川峨眉山Alt.2400m
♂外部生殖器のプロポーションや末端構造に於いて(あるいは翅の概形や斑紋・色彩などに於いても)、クロキマダラヒカゲNeope agrestisと対極的な印象を持つのが、タイワンキマダラヒカゲ台湾亜種(N.bremeri watanabei)である。N.bremeriの大陸産原名亜種と台湾亜種の間には、一般的常識による種間差を優に越える差があるように見受けられるが、形質の一定方向への“傾き”であると捉えれば、両者を同一種として扱うことは妥当であると思われる。ただしその場合、反対方向の傾斜上に位置するN.agrestisの処遇も考慮しなければならない。大陸東部(杭州)産N.bremeri原名亜種は、全体の傾斜の中で、台湾亜種N.bremeri watanabeiと、大陸西南部高地産 N.agrestisとの中間に位置する、と言ってよい(日本産のN.niphonicaについてもそれに近いことが言えそうである)。
四川成都市近郊(青城山)産もN.bremeriに含めた。杭州市産と同じくbremeriの原名亜種に置く事が妥当か否かについては今後の詳しい検証を待たねばならないが、盛夏に同一種の夏型(外部生殖器形状の一致を検証済み)が出現することから、夏型の出現しないN.agrestisとは一線を区し、(青城山に隣接した西嶺雪山で撮影した夏型♀共々)狭義のbremeriに含めておく。
ほかに青城山からは、性票(特殊鱗の集中散布)を欠く春型♂を記録していて、「中国のチョウ」では暫定的にN.pulahoidesとしておいた。N.pulahoidesやN.pulahinaは、広義にはN.pulahaの一群に所属するものと考えられるが、この個体は、あるいはpulaha ではなくbremeriに繋がるのかも知れない。先に記したように、性標(特殊鱗の有無や散布範囲)が種間関係に於いてどのような意味を持ち、分類指標としてどの程度の重要性を示すのかを検証することは、今後の重要な検証課題であると思われる。
それらの課題はさておき、(N.niphonicaの分類上の処遇はともかく)N.bremeri, N.agrestis, N.niphonica等の各種が、同一の形質傾斜上に置かれた、ひとつの上種super-speciesを形成する可能性を、一応の結論としておきたい。
では、サトキマダラヒカゲNeope goschevitschiiは、どの傾斜上に位置するのであろうか? 筆者の結論は、少々乱暴ではあるが「アリサンキマダラヒカゲN.pulaha」が、国外に於ける日本固有種「サトキマダラヒカゲ」の対応種であるということ。
“乱暴”の実態を補足しつつ、中国西南部に於ける「サトキマダラヒカゲ」対応集団の写真を紹介していく。
Neope pulaha【仮同定】(手前)とN.agrestis(奥)2012.6. 29 雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】 2013.5. 7 雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】 2013.5. 8 雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】 2013.5. 8 雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】 2013.5. 8 雲南梅里雪山Alt.2300m(上と同一個体)
Neope pulaha【仮同定】 2013.5. 10 雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m
Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m
Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m
Neope pulaha【仮同定】 2009.6. 11雲南梅里雪山Alt.3500m
Neope pulaha【仮同定】 2009.6. 11雲南梅里雪山Alt.3500m
Neope pulaha【仮同定】 2012.6. 30雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】 2012.6. 29雲南梅里雪山Alt.2300m(12枚前と同一カット)
現時点では(比較的限られた地域で広範囲とはいえないが)少なくとも4つの地域に於いて“典型Neope”と呼ぶべき2種の混棲が知られている。先にも述べたように、重要なことは、単に2種が混棲していると言うことよりも、それらの地域に見られる(一見多様な変異形質を示す)“典型Neope”の全てが、そのどちらかの種に帰属する、ということである。
ひとつはもちろん日本産の2種。そして台湾のタイワンキマダラヒカゲ台湾亜種Neope bremeri watanabeiとアリサンキマダラヒカゲ台湾亜種Neope pulaha didia。以下に述べる雲南省北部山地に於けるクロキマダラヒカゲNeope agrestisとNeope pulaha類似種。およびそれと同一の組み合わせと考え得るミャンマー北部産の2種(Butterflies Musiam展示の過程で多数の標本を検証)。それ以外の地域では、今のところ前者(bremeriなどniponica対応集団)のみしか確認していないが、より多くの地域で“2種”が混棲している可能性も有り得る。
先に検証したように、niphonica~bremeri~agrestisは、同じ傾斜上に位置するniphonica対応集団と考えられる。ならば、一方のgoschchevitschii~pulaha didia(台湾亜種)~大陸東南部産pulaha類似種も、同じ傾斜上に置かれる対応集団、と考えるのは早計に過ぎるであろうか?
常識的には、暴論には違いない。アリサンキマダラヒカゲ台湾亜種N.pulaha didiaは(中国大陸産アリサンキマダラヒカゲN.pulahaのいくつかの亜種ともども)、外観上、様々な点でサトキマダラヒカゲN.goschevitschiiとは対極的な特徴を示している。 ヤマとサトの有意な区別点たる、「前翅外縁のカーブの仕方」「前翅表第0室眼状紋内の黒点の位置」「後翅裏面基部の3個の丸紋の並び方」など、いずれもサトとの共通性は全く示さず、むしろずっとヤマ的だ。アリサンとサトは、“典型Neope”の中では最も離れた位置にあると考えるのが妥当であろう。
大陸西南部高地産のN.pulaha類似種に於いても、以下に述べる斑紋などの幾つかの特徴はともかく、より広域に分布するN.bremeri~N.agrestisに対して、明らかに(高標高地のみに)分布域が限られている。日本のヤマとサトの関係とは正反対なのである。
しかし、日本と中国では、近縁種間の組み合わせに於いて生態的地位が逆転する、という現象は、枚挙にいとまない。また、表現された形質が、必ずしも系統を反映しているとは限らない、ということも、多くの例で知られるところである。
そして、非常に気になる事実がひとつ。最初に示した、ヤマとサトの、最も安定的な分類指標である「③後翅裏面基部の3つの丸紋の並び方」に於いて、大陸西南部(雲南省北部)山地産のN.pulaha類似種と、日本産N.goschevitschiiは、見事に一致するのである。
そのような前提で改めてこのN.pulaha類似種に注目すれば、裏面の色調や全体的な印象も、どこかサトに共通する印象が感じられる(ただし現地での観察に於いて、ことに飛翔時には本種は真っ黒に見え、一方、筆者が「中国のチョウ」で“クロキマダラヒカゲ”と名付けたNeope agrestisは、遥かに明るい色調~赤褐色~に見えることから、和名については再検討するべきと考えている)。
ちなみに、最初に記したごとく、♂外部生殖器の形状は、ヤマとサトの間で酷似し、全く相同のようにも見えるが、川副昭人氏も指摘し(若林守男氏との共著による「日本産蝶類大図鑑」保育社) 筆者も「中国のチョウ」で解説したように、非常にデリケートではあるが、それなりの安定差が見出される。すなわち、ヤマではpenisに生じる鋸歯群の個々の鋸歯が相対的に大型で、やや疎らに散布される傾向があるのに対し、サトでは個々の鋸歯がより小型で、大きさや散布密度が一定しているということ。また、(あくまで傾向的・印象的な相違に過ぎぬが)uncus概形の全体的な印象が、サトでは茫洋とした感じで、ヤマのような引き締まりに欠ける気がする。それらの傾向は、buremeri watanabei~bremeri buremeri~agrestisと、pulaha complexの間にも当て嵌まるように思える。
筆者は、原則、Neope分布圏内の多くの地域で、2組の“典型Neope”の組み合わせが成されているのではないかと考えている。そして、どの組み合わせの2つの種も、ヤマ、サト、それぞれの傾斜上に位置しているのではないかと。日本列島に於ける2つの種の酷似は、並行的に成された、2次的な類似だと解釈したい。
むろん、それぞれの組み合わせに於ける2種が、同じ傾斜上の同一種群間に帰属するのではなく、各地での組み合わせごとに、(少なくとも“サト”に対応する側は)別系統の集団からなっている可能性も否定出来ない。ここに示した雲南省北部山地産“アリサンキマダラヒカゲNeope pulaha類似種”が、果たして系統上、(台湾産亜種をはじめとした)ほかのアリサンキマダラヒカゲ各亜種に繋がるのかなど、全く分かっていない。
それらを含めた、より詳しい検証が成されなければならない。幼生期、生態、行動、生理の比較、性標(特殊鱗粉)の散布状況と機質的機能的意味の解明、むろんDNA解析による系統的考察。
頼りとするのが高橋真弓氏であることは、今も昔と変わらない。氏は今も(国内外に調査旅行に赴かれるなど)積極的に活動を続けていられるが、来年には古希(80歳)を迎えられる。高橋氏に全てを頼るというのは酷であろう。といって、小生(青山)や日本に於けるLethe族研究の第一人者である鈴木英文氏も、前期高齢者(65歳)に突入する。我々が成すことの出来る範囲は限られていよう。若い方々(中国をはじめとする国外の人々を含む)によるアグレッシブな探求を、切に願うところである。
Neope agrestis 2012.6. 29雲南梅里雪山Alt.2300m
Neope agrestis 2009.6. 1雲南香格里拉-虎跳峡Alt.2800m
Neope agrestis 2009.6. 1雲南香格里拉-虎跳峡Alt.2800m
Neope agrestis 2013.5. 9雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m
Neope pulaha【仮同定】 2009.6. 11 雲南梅里雪山Alt.3500m
Neope pulaha【仮同定】 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m
Neope pulaha【仮同定】 2013.5. 10 雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】(下4頭と上右)/Neope agrestis(上左2頭) 2009.6. 1 雲南香格里拉-虎跳峡Alt.2800m
Neope pulaha【仮同定】(左)/Neope agrestis(右) 2013.5. 10 雲南梅里雪山Alt.2300m
Neope pulaha【仮同定】(左)/Neope agrestis(右)
追記
小生はコレクションの趣味を全く持たず、原則としていかなる地域でも採集は行わないが、Neopeをはじめ、Ypthima(ウラナミジャノメ属)、 Pieris(モンシロチョウ属)、 Parnara(イチモンジセセリ属)等々、ごく地味ではあるが、日本やアジアの蝶相(ひいては各地域のアイデンティティイ)の解明に当たって極めて重要な意味をもつ幾つかの分類群については、必要に応じて採集を行っている。
注1:大抵の場合、補虫網を使わず手で摘まんだり叩き落したり足で踏んづけたりして採取するので、コレクターの視点からの標本的価値は全くない。
注2:それらの標本は中国各地の博物館や来年度にオープン予定の「静岡県立博物館」などに保管。
上左:goschevvitschiiとniphonica
上右:bremeri bremeriとbremeri watanabei
下左:agrestis
下右:pulahaほか
【「中国のチョウ」より】
・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・・
僕のブログのうち、いわば“趣味”でアップしている「ElvisとBeatlesの狭間で~Johnny Tillotsonの時代」や「朝と夜の狭間で~My Sentimental Journey」と異なり、昆虫や植物関係の話題については、“仕事”の一環と考えています。「青山潤三ネイチャークラブ」の会員諸氏や、それに準じる協力者諸氏に向けての発信です。(たまたま訪問された方はともかく)継続してブログを訪問される方には、各自何らかの形での自主的な援助を望んでいる次第です。その旨、ご承知おき下さい。





