がんの予防や治療における漢方治療の存在意義を考察しています。がん治療に役立つ情報も紹介しています。
「漢方がん治療」を考える
カレンダー
| 2025年1月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | |||||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| 12 | 13 | 14 | 15 | 16 | 17 | 18 | ||
| 19 | 20 | 21 | 22 | 23 | 24 | 25 | ||
| 26 | 27 | 28 | 29 | 30 | 31 | |||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
736)mTORC1活性の阻害をターゲットにしたがん治療

図:インスリンやインスリン様成長因子-1や成長ホルモンなどの増殖因子が細胞の受容体に作用するとPI3キナーゼ(PI3K)というリン酸化酵素が活性化され、これがAktというセリン・スレオニンリン酸化酵素をリン酸化して活性化する(①)。活性化したAktはTSC1/TSC2を阻害してRheb(Ras homolog enriched in brain)を活性化し、mTORC1を活性化する(②)。AktはmTORC1を直接活性化する作用もある(③)。メトホルミン(④)とビタミンD3(⑤)はAMP依存性プロテインキナーゼ(AMPK)を活性化する(⑥)。活性化したAMPKは、TSC1/TSC2の活性化(⑦)やmTORC1の直接的な抑制害作用(⑧)によってmTORC1活性を抑制する。ラパマイシンはFKBP12(12kDa FK506 binding protein)と結合し、mTORC1の活性を阻害する(⑨)。オーラノフィンはPI3K/Akt/mTORC1経路を阻害する(⑩)。メイベルメクチンはPAK-1阻害作用によってPI3K/Akt/mTORC1経路を阻害する(⑪)。イトラコナゾールは複数のメカニズムでmTORC1活性を抑制する(⑫)。したがって、これらを組み合せるとTORC1の活性を強く抑制し、がん細胞の増殖を抑制できる。
736)mTORC1活性の阻害をターゲットにしたがん治療
【がん細胞が増殖するには栄養とエネルギーが必要】
正常細胞は増殖や細胞死が厳密に制御されていて、無制限に増殖することはありません。
がん細胞は、遺伝子の変異や発現異常によって細胞周期やアポトーシス(細胞死)の制御に異常が起こって、無制限な自律増殖能を獲得しています。
がん細胞は栄養が十分に供給されれば無制限に増殖できます。増殖を促進するために、がん細胞はグルコースやアミノ酸の取込みと消費が増加しています。
がん細胞が増殖するための2大栄養素はグルコースとグルタミンです。がん細胞では、グルコースとグルタミンの取込みが増えています。細胞の増殖を維持するために、物質合成とエネルギー産生のための栄養源が必要だからです。

図:増殖活性の高いがん細胞はグルコースとグルタミンの取込みと代謝が亢進している。
栄養やエネルギーの供給が減少すれば増殖速度は低下し、栄養やエネルギーの供給が停止すれば細胞は増殖できません。
栄養源の取込みを促進し、細胞の増殖を亢進する働きを担っているのが、哺乳類ラパマイシン標的タンパク質複合体1(mTORC1)です。mTORC1の活性を阻害すれば、がん細胞の増殖は抑制できます。
【ラパマイシンは哺乳類ラパマイシン標的タンパク質複合体1(mTORC1)を阻害する】
ラパマイシン(Rapamycin)は1970年代にイースター島(モアイ像で有名な南太平洋の島)の土壌から発見されたStreptomyces hygroscopicsという放線菌の一種が産生する有機化合物です。シロリムス(Sirolimus)という別名で呼ばれることもあります。
ラパマイシンは免疫抑制作用があり、米国では臓器移植の際の拒絶反応を防ぐために使用されています。さらに、平滑筋細胞増殖抑制作用や抗がん作用や寿命延長効果が知られています。
平滑筋細胞増殖抑制作用に関しては、狭心症や心筋梗塞の治療に使われる血管内ステントに冠動脈再狭窄予防効果を目的としてラパマイシンを配合したステントが製品化され、心臓カテーテル治療において使用されています。また、リンパ脈管筋腫症の治療薬としても使用されています。
ラパマイシンの生体内のターゲット分子が、哺乳類ラパマイシン標的タンパク質(mammalian target of rapamycin)、略してmTOR(エムトール)というタンパク質です。 mTORはラパマイシンの標的分子として同定されたセリン・スレオニンキナーゼ(タンパク質のセリンやスレオニンをリン酸化する酵素)で、細胞の分裂や生存などの調節に中心的な役割を果たすと考えられています。
酵母におけるラパマイシンの標的タンパク質が見出されてTOR(target of rapamycin)と命名され、後に哺乳類のホモログ(相同体)が見出されてmTOR(哺乳類ラパマイシン標的タンパク質)と命名されました。
mTORにはmTOR複合体1(mTORC1)とmTOR複合体2(mTORC2)の2種類があります。mTORに幾つかの他のタンパク質が結合して複合体を形成しますが、結合しているタンパク質の違いで2種類の複合体ができ、異なる機能を担っています。
mTORC1は成長因子や、糖やアミノ酸などを含む栄養素のセンサーとして機能し、mTORC2は細胞骨格やシグナル伝達の制御をしています。
インスリンやインスリン様成長因子(IGF-1)によって活性化されるのはmTORC1の方です。ラパマイシンで阻害されるのもmTORC1の方です。
ラパマイシンは単独でTOR 活性を阻害するのではなく、細胞内でFKBP12(12kDa FK506 binding protein)と結合し,このFKBP12-ラパマイシン複合体がTOR に結合してTOR 活性を阻害します。

図:哺乳類ラパマイシン標的タンパク質複合体1(mTORC1)は複数のタンパク質から構成されるセリン・スレオニン・リン酸化酵素で、インスリンやインスリン様成長因子-1(IGF-1)や上皮成長因子(EGF)や血小板由来増殖因子(PDGF)などの増殖因子によって活性化される。
mTORC1はタンパク質翻訳の開始因子であるelF4Eを抑制する4E結合タンパク質(4E-BP1)をリン酸化してその機能を抑制する。また、リボソームの生合成を促進するS6K1をリン酸化して活性化する。これらの作用によってmTORC1はタンパク質合成を促進する。その他、多くの標的タンパク質をリン酸化することによって脂質や核酸の合成を亢進し、細胞内小器官の消化・再利用に重要なオートファジーを抑制する。ラパマイシンはFKBP12と結合し、mTORとraptorの相互作用を阻害することによってmTORC1の活性を阻害する。
【mTORC1とAMPKは栄養と増殖シグナルとエネルギー量を感知して増殖を制御する】
細胞の増殖というのは、栄養とエネルギーが利用できる状態にあるときに、新たな細胞構成成分(タンパク質、核酸、脂質など)を合成して、細胞の数を増やす生化学的プロセスのことです。
したがって、増殖するためには、細胞を新たに作る材料(栄養素)とエネルギー(糖質や脂質を酸化して得られるATP)が必要です。
増殖因子や成長因子やホルモンなどによって細胞増殖の指令(シグナル)が来たときに、栄養素とエネルギーの供給が十分にある条件で、タンパク質や脂質の合成を促進して細胞増殖を実行するのが哺乳類ラパマイシン標的タンパク質複合体1(mTORC1)です。
インスリン、インスリン様成長因子-1(IGF-1)、成長ホルモンなどの増殖刺激が細胞に作用すると、それらの受容体を介してPI3キナーゼ(PI3K)というリン酸化酵素が活性化され、これがAktというセリン・スレオニンリン酸化酵素をリン酸化して活性化します。

図:PI3キナーゼ-AKT経路は,細胞外からのシグナルを細胞内に伝える主要な経路の一つで、増殖因子など様々な刺激により活性化され、細胞の生存と増殖・大きさの制御、細胞運動、代謝の制御など多くの細胞機能に関与している。ホスファチジルイノシトール3-キナーゼ(PI3K)は,細胞膜の構成成分であるイノシトールリン脂質をリン酸化する酵素で,産生したPI3,4,5-三リン酸(PIP3)がAktをリン酸化する。Aktは多くのがん組織で活性しておりAKTシグナル伝達系は、細胞増殖や生存、細胞サイズや栄養素利用への応答性、グルコース代謝、組織浸潤および血管新生など多くの細胞プロセスを制御している。がん細胞の生存と増殖はAKTシグナル伝達系の活性に依存度が高いので、AKTシグナル伝達系の阻害はがん細胞の増殖抑制やアポトーシス誘導を引き起こす。
ホスファチジルイノシトール3-キナーゼ(PI3K)は,細胞膜の構成成分であるイノシトールリン脂質をリン酸化する酵素です。ホスファチジルイノシトール4,5-二リン酸(PIP2)の3位のOHをリン酸化してホスファチジルイノシトール3,4,5-三リン酸(PIP3)を生成する酵素です。PIP3がAktをリン酸化して活性化します。
PTENは脱リン酸化する酵素でPIP3をPIP2に変化することによってAktの活性化を阻止します。

図:ホスファチジルイノシトール3-キナーゼ(PI3K)は,細胞膜の構成成分であるイノシトールリン脂質をリン酸化する酵素で,産生したPI3,4,5-三リン酸(PIP3)がAktをリン酸化する。Aktは多くのがん組織で活性しておりAKTシグナル伝達系は、細胞増殖や生存、細胞サイズや栄養素利用への応答性、グルコース代謝、組織浸潤および血管新生など多くの細胞プロセスを制御している。PTENはPIP3を脱リン酸化してPIP2に変換し、Akt活性化を阻止する。
活性化したAktは、細胞内のシグナル伝達に関与する様々な蛋白質の活性を調節することによって細胞の増殖や生存(あるいは死)の調節を行います。このAktのターゲットの一つがmTORC1です。
AKTは結節性硬化症(Tuberous Sclerosis:TSC)の原因遺伝子産物であるTSC1/TSC2複合体のTSC2をリン酸化して活性を低下させ、低分子 GTPaseの一種である Rheb (Ras Homolog enriched in Brain)の活性を高めます。活性化したRhebは mTOR のキナーゼ触媒ドメインに直接結合してmTORC1のキナーゼ活性を高めます。
アミノ酸は別の経路でmTORC1の活性を高めます。mTORC1の活性化においてアミノ酸(哺乳類では特にロイシン)が必須であり、これはアミノ酸が十分である場合に限り細胞がタンパク合成を開始できる仕組みと言えます。
牛乳タンパク質はロイシンの含有量が多いので、mTORC1を活性化する作用が強いので、成長を促進する効果が高い食品となります。
このようにして、栄養源と増殖シグナルを感知して細胞の成長や分裂を促進するのがmTORC1です。
一方、細胞内のエネルギー低下を感知するのがAMP依存性プロテインキナーゼ (AMPK)です。グルコース欠乏や低酸素などにより細胞内ATP 量が減少すると、AMP/ATP 比の増加に伴いAMPKが活性化されます。
AMPKはmTORC1活性を抑制して、異化作用の亢進や細胞成長の停止をもたらし、エネルギー消費の抑制(同化抑制)とエネルギー産生の亢進(異化促進)へと細胞の代謝をシフトさせます。

図:AMPKは触媒作用を持つαサブユニットと、調節作用を持つβサブユットとγサブユニットから構成されるヘテロ三量体として存在する(①)。運動やカロリー制限や虚血や低酸素などによってATPが減少してAMP/ATP比が上昇すると(②)、γサブユニットに結合していたATPがAMPに置換する(③)。これによってAMPKの構造変化が起こると、LKB1というリン酸化酵素の親和性が高まり、αサブユニットのスレオニン172がリン酸化されると、さらにAMPKの活性が高まる(④)。活性化したAMPKは異化を亢進してエネルギー産生を亢進し、物質合成(同化)を抑制するように代謝をシフトする(⑤)。活性化したAMPKはmTORC1(哺乳類ラパマイシン標的タンパク質複合体1)を阻害し、がん細胞の増殖抑制や、抗老化や寿命延長の効果を引き起こす(⑥)。
AMPKは運動やカロリー制限の他、メトホルミンやビタミンD3で活性化できます(下図)。

図:AMPKの172番目のスレオニン(Thr-172)がLKB1でリン酸化されると(①)、AMPKは最大に活性化される(②)。運動やカロリー制限はATPが減少してAMP/ATP比を上昇してAMPKを活性化する(③)。メトホルミンはミトコンドリアの呼吸鎖を阻害してATP産生を低下させる機序とLKB1を活性化する両方の機序でAMPKを活性化する(④)。ビタミンD3は細胞内のフリーのカルシウムを増加させ、カルモジュリンキナーゼキナーゼβ (CaMKKβ)を活性化させてAMPK活性を亢進する(⑤)。
【オーラノフィンはPI3K/AKT/mTOR経路を阻害する】
オーラノフィン(Auranofin)は、関節リュウマチにおける炎症反応や免疫異常を抑制して寛解へと導く経口金製剤として1985年以降臨床で使用されています。炎症細胞の機能抑制や、免疫細胞に作用して自己抗体の産生を抑制して、関節における炎症を抑制します。
最近、オーラノフィンの抗腫瘍効果が注目されています。米国ではがん治療へのオーラノフィンの効果を検討する第2相臨床試験の実施がFDA(食品医薬品局)から承認されています。
今まで報告されたオーラノフィンの抗がん作用のメカニズムは多様です。DNAやRNAやタンパク質の合成阻害、ミトコンドリアのチオレドキシン還元酵素やグルタチオン-S-トランスフェラーゼやプロテアソームの機能阻害、抗炎症作用(IL-6/STAT3経路の阻害、NF-κB活性化の阻害など)、ヒストン・アセチル化亢進など多くの作用機序が報告されています(オーラノフィンの抗がん作用の詳細はこちらへ)。
オーラノフィンがPI3K/AKT/mTOR経路を阻害して抗がん作用を発揮するメカニズムも報告されています。以下のような報告があります。
Auranofin-mediated inhibition of PI3K/AKT/mTOR axis and anticancer activity in non-small cell lung cancer cells(非小細胞性肺がんにおけるオーラノフィンによるPI3K/AKT/mTOR経路の阻害と抗腫瘍活性)Oncotarget. 2016 Jan 19; 7(3): 3548–3558.
【要旨】
オーラノフィンは慢性関節リュウマチの治療に使われている金複合体で、安全性が高く、さらに近年、白血病や固形がんに対する治療効果が検討されている。しかしながら、肺がんに対するオーラノフィン単独の抗腫瘍効果については十分に検討されていない。
肺がんに対してオーラノフィンが単独で抗腫瘍活性を示すかどうかを検討する目的で、10種類の非小細胞性肺がん細胞株を用いてオーラノフィンの抗腫瘍活性を検討した。
その結果、オーラノフィンは非小細胞性肺がん細胞に対して、増殖抑制効果を示し、その50%増殖阻止濃度(IC50)は最大で1.0μMであった。
オーラノフィンに感受性の肺がん細胞に対して、オーラノフィンは細胞死(アポトーシス)を誘導した。
さらに、肺がん細胞のオーラノフィンに対する感受性はTXNRD1発現と逆相関を認めた。
TXNRD1を発現するプラスミドを一過性に発現させると、オーラノフィン感受性のCalu3細胞はオーラノフィンに抵抗性を示すように変化した。これはTXNRD1の高発現がオーラノフィンに対する抵抗性の因子の一つであることを示している。
さらにメカニズムの検討によって、S6、 4EBP1、 Rictor、 p70S6K、 mTOR、 TSC2、 AKT 、GSK3などを含むPI3K/AKT/mTOR経路の多くのタンパク質の発現とリン酸化を阻害することが明らかになった。
このようなオーラノフィンによるPI3K/AKT/mTOR経路の阻害はTXNRD1の一過性の発現によって阻止された。これはPI3K/AKT/mTOR経路の制御にTXNRD1が関与している可能性を示唆している。
肺がんを移植したマウスにオーラノフィンを投与すると明らかな毒性(副作用)を示すことなく、腫瘍の増大が顕著に抑制された。これらの結果は、肺がん治療にオーラノフィンを使用する根拠を提示している。
TXNRD1というのはチオレドキシン還元酵素1(Thioredoxin reductase 1)のことです。
オーラノフィンはチオレドキシン還元酵素を阻害する作用があります。
チオレドキシン還元酵素はPI3K/AKT/mTOR経路の制御(活性化)に関与しており、チオレドキシン還元酵素の阻害剤のオーラノフィンがPI3K/AKT/mTOR経路を阻害するというメカニズムです。
【ビタミンD3は複数のメカニズムでmTORC1の活性を阻害する】
ビタミンDは、血清カルシウム濃度の恒常性や骨代謝における作用が主な働きだと考えられていますが、最近の研究によって、細胞の増殖や分化や細胞死(アポトーシス)や免疫機能など多彩な生理活性の制御に重要な役割を担っていることが明らかになっています。
ビタミンD3がmTORC1(mammalian target of rapamycin complex 1:哺乳類ラパマイシン標的蛋白質複合体1)の活性を抑制する作用も報告されています。
前述のように、ビタミンD3はカルモジュリンキナーゼキナーゼβ (CaMKKβ)を活性化させてAMPK活性を亢進するメカニズムが報告されています。AMPKはmTORC1活性を阻害します。
さらに、活性型ビタミンD3(1,25(OH)2ビタミンD3)がビタミンD受容体を介するメカニズムでDDIT4(DNA-damage-inducible transcript 4)という遺伝子の発現を亢進し、このDDIT4がTuberous Sclerosis Complex 1/2(TSC1/2)を活性化し、これがRheb(Ras homolog enriched in the brain)を阻害してmTORC1の活性を抑制するという作用機序が報告されています。(下図)

図:インスリンやインスリン様成長因子-1(IGF-1)などの増殖刺激はPI3K/Akt/mTORC1シグナル伝達系を活性化する(①)。ビタミンD3はビタミンD受容体を介する遺伝子転写活性によってDDIT4(DNA-damage-inducible transcript 4)タンパク質の発現を亢進する。このDDIT4はTuberous Sclerosis Complex 1/2(TSC1/2)を活性化し、これがRheb (Ras homolog enriched in the brain)を阻害してmTORC1の活性を抑制する(②)。ビタミンD3はBeclin 1の発現を亢進する。Beclin 1はPI3キナーゼ(PI3K)と相互作用してmTORC1の働きを阻害し、オートファジーを亢進する(③)。ビタミンD3は細胞内のフリーのカルシウム濃度を高めてカルモジュリンキナーゼキナーゼβ (CaMKKβ)を活性化しAMP活性化プロテイン・キナーゼ(AMPK)活性を亢進する(④)。活性化したAMPKはTSC1/TSC2を活性化し、Rhebを阻害してmTORC1の活性を抑制する(⑤)。このようにビタミンD3は複数のメカニズムでmTORC1活性を抑制する。
メトホルミンとオーラノフィンとビタミンD3は非常に安価で安全性が高く、様々なメカニズムで抗がん作用を示します。
メトホルミンやビタミンD3やオーラノフィンは「医薬品の再開発(Drug Repositioning)」として、がん治療にもっと利用されて良いと思います。
【コレステロールの細胞内輸送の阻害はmTORシグナル伝達系と血管新生を阻害する】
最近、コレステロールの細胞内輸送の阻害をターゲットにしたがん治療法が注目されています。
コレステロールの細胞内輸送、つまり細胞内利用を阻害するとmTORシグナル伝達系や血管新生を阻害できるというメカニズムが提唱されています。
以下のような総説論文があります。
Cholesterol Trafficking: An Emerging Therapeutic Target for Angiogenesis and Cancer(コレステロールの細胞内輸送:血管新生とがんの新たな治療標的)Cells. 2019 Apr 28;8(5):389.
【要旨】
コレステロールは細胞膜の必須の構造成分である。 構造的な役割に加えて、さまざまなステロイドホルモンの前駆体として機能し、細胞内シグナル伝達においても多様な機能を持っている。
細胞シグナル伝達における機能の1つとして、コレステロールが血管新生の調節に重要な役割を果たしていることが多く研究で明らかになっている。 このレビューでは、血管新生におけるコレステロールの役割に関して、特に、内皮細胞のシグナル伝達におけるコレステロールの細胞内輸送に重点を置いて解説する。 コレステロール輸送の小分子阻害剤と、血管新生およびがんを標的とするそれらの前臨床および臨床開発についても説明する。
コレステロールは細胞膜や様々な細胞小器官の脂質二重膜を構成する成分の1つです。血液中のコレステロールは、細胞の受容体を介して細胞内に取り込まれた後、リソソームまで運ばれ、リソソームの酵素によって遊離コレステロールが細胞質へ放出され、脂質二重膜の構成成分として利用されたり、ホルモンの原料となったり、脂肪滴として細胞内に貯蓄されたりします。
このコレステロールの細胞内輸送が阻害されると、細胞内シグナル伝達系に異常が起こるということです。がん細胞の増殖阻止や血管新生阻害にも関連します。

図:血液中の低密度リポタンパク質(LDL)はコレステロールをリン脂質が包み込んでいる(①)。細胞膜のLDL受容体を介してLDLはエンドサイトーシスを介して細胞内に取り込まれる(②)。LDL受容体は細胞膜に戻って再利用される(③)。エンドソームはリソソームと癒合し(④)、リソソームの酵素によって遊離コレステロールが細胞質へ放出される(⑤)。
以下のような報告があります。
Astemizole Inhibits mTOR Signaling and Angiogenesis by Blocking Cholesterol Trafficking(アステミゾールはコレステロールの細胞内輸送を阻止することによってmTORシグナル伝達系と血管新生を阻害する)Int J Biol Sci. 2018 Jun 23;14(10):1175-1185
【要旨】
コレステロールは、内皮細胞の膜タンパク質機能とシグナル伝達において重要な役割を果たしている。したがって、コレステロールの細胞内輸送を阻害することは、血管新生を阻害するための効果的な方法となる。
最近、我々は、抗ヒスタミン薬であるアステミゾール(astemizole)がコレステロールの細胞内輸送の阻害剤であることを発見した。
この研究では、アステミゾールがコレステロール輸送に関与するリソソーム表面タンパク質であるニーマンピック病C1型(NPC1)タンパク質のステロール感知ドメインに結合することにより、リソソームにコレステロールの蓄積を誘導することを発見した。
アステミゾールによるコレステロール輸送の阻害は、膜コレステロールの枯渇につながり、SREBP1の核局在化を引き起こした。
膜コレステロールの枯渇は、リソソーム表面からの哺乳類ラパマイシン標的タンパク質(mTOR)の解離と、mTORシグナル伝達の不活性化をもたらした。これらの影響は、外因性コレステロールの添加によって効果的に救済された。
アステミゾールは、コレステロール依存的に内皮細胞の増殖、遊走、管形成を抑制した。さらに、アステミゾールはコレステロール依存的にゼブラフィッシュの血管新生を阻害した。
以上の実験結果は、アステミゾールが内皮細胞のコレステロール輸送と血管新生を阻害する新しいクラスのNPC1アンタゴニストであることを示唆している。
アステミゾール(astemizole)は過去に抗ヒスタミン薬として使用されていましたが、副作用の問題で販売中止になり、1999年に世界的に市場から撤退しています。したがって、現在では医薬品としては使用されていません。
SREBP1(Sterol regulatory element-binding protein 1)は脂質生合成の重要な転写調節因子です。インスリン刺激を受けると、SREBP1は小胞体からゴルジ体へ輸送され、そこでプロセシングを受けてから、核へと輸送されコレステロールや脂肪酸の合成に関わる遺伝子を誘導します。「SREBP1の核局在化を引き起こす」というのは、脂質生合成が誘導されることを意味します。
コレステロールの細胞内輸送を阻止することは、mTORシグナル伝達系と血管新生を阻害することによってがん細胞の増殖を抑制することを示唆しています。
ニーマン・ピック(Niemann-Pick)病という先天性代謝異常症があります。
酸性スフィンゴミエリナーゼが欠損するA型、B型とNPC1またはNPC2蛋白の異常によって起こるC型に分類されます。いずれも常染色体劣性遺伝形式を示す遺伝病です。肝臓、脾臓、骨髄の網内系細胞と神経細胞にスフィンゴミエリン、コレステロール、糖脂質などが蓄積します。
この論文で出ている「ニーマンピック病のC1型(NPC1)タンパク質」というのはC型のニーマン・ピック病で異常を起こしているタンパク質です。NPC1遺伝子の変異によってコレステロールが蓄積します。
前述のようにLDL(低密度リポタンパク質)はLDL受容体を介して肝臓などの細胞中にエンドサイトーシスという方法で吸収されます。吸収されたLDLはエンドソームという小胞を形成し、この小胞はリソソーム内の酵素によって分解されます。NPC1タンパク質はリソソームからコレステロールを排出するときに必要なタンパク質で、NPC1遺伝子に変異があるニーマン・ピック病C1型では、コレステロールが細胞質内に蓄積して発症します。
この論文では「アステミゾールがコレステロール輸送に関与するリソソーム表面タンパク質であるニーマンピック病のC1型(NPC1)タンパク質のステロール感知ドメインに結合することにより、リソソームにコレステロールの蓄積を誘導した」という結果です。 
図:リソソーム内に輸送されたLDL(低密度コレステロール)はリソソームの酸性リパーゼ(lysosomal acid lipase)によって加水分解されて遊離コレステロールとなる(①)。コレステロールは、リソソーム内腔のニーマンピック病C型(NPC)NPC2に結合し(②)、リソソームの内膜側にあるNPC1のN末端ドメイン(N-terminal domain:NTD)に転移する(③)。コレステロールはNPC1のN末端ドメイン(NTD)からステロール感知ドメイン(sterol-sensing domain :SSD)に転送され(④)、リソソームから細胞質に輸送される(⑤)。アステミゾールはNPC1のステロール感知ドメイン(SSD)に結合し、NPC1によるコレステロール輸送を妨害する(⑥)。その結果、リソソームでのコレステロールの蓄積と細胞質の細胞内コンパートメントでのコレステロールの枯渇が誘導される。
【イトラコナゾールはコレステロールの細胞内輸送を阻害する】
イトラコナゾール(商品名イトリゾール等)は、水虫等の白癬(はくせん)菌症、口腔や食道カンジダ症等に広く使用されています真菌(カビなど)治療薬です。真菌コレステロールを形成する主な酵素を阻害して真菌を死滅します。

イトラコナゾールには血管新生阻害作用が報告されています。さらに、ヘッジホッグシグナル伝達系の阻害作用、オートファジー誘導によるがん細胞の細胞死誘導作用などの報告があり、がんの代替治療薬としても注目を集めています。
例えば、以下のような報告があります。
Itraconazole exerts its anti-melanoma effect by suppressing Hedgehog, Wnt, and PI3K/mTOR signaling pathways(イトラコナゾールは、ヘッジホッグ、Wnt、およびPI3K / mTORシグナル伝達経路を抑制することにより抗メラノーマ効果を発揮する)Oncotarget. 2017 Apr 25;8(17):28510-28525.
【要旨の抜粋】
広く使用されている全身性抗真菌薬であるイトラコナゾールは、基底細胞がん、前立腺がん、および非小細胞肺がんに対する抗腫瘍効果について検討されている。
この研究の目的は、悪性黒色腫に対するイトラコナゾールの効果を検討し、その作用メカニズムの詳細を明らかにすることである。
In vivoの異種移植腫瘍マウスモデルにおいて、イトラコナゾールが、イトラコナゾール非投与マウスと比較して、黒色腫細胞の増殖を阻害し、黒色腫細胞移植マウスの生存を延長することを認めた。
また、イトラコナゾールで処理された腫瘍組織ではKi-67陽性細胞の減少が認められ、イトラコナゾールが細胞増殖を有意に阻害する可能性を示唆した。
in vitroの実験系で、イトラコナゾールはSK-MEL-28とA375の両方のヒトメラノーマ細胞の増殖とコロニー形成を阻害した。
さらに、イトラコナゾールはGli-1、Gli-2、Wnt3A、β-カテニン、およびサイクリンD1の発現を有意に抑制する一方で、Gli-3およびAxin-1の発現を亢進した。これは、ヘッジホッグ・シグナル伝達系とWntシグナル伝達経路に対するイトラコナゾールの強力な阻害効果を示している。
さらに、イトラコナゾールは、PI3K / mTORシグナル伝達経路(p70S6K、4E-BP1、AKTのリン酸化の抑制によって示される)を大幅に抑制するが、MEKまたはERKのリン酸化には影響しない。
我々のデータは、イトラコナゾールが、ヘッジホッグ、Wnt、およびPI3K / mTORシグナル伝達経路の阻害を介して悪性黒色腫細胞の増殖を阻害することを示唆している。
これらの結果は、イトラコナゾールがいくつかの強力な抗メラノーマ作用を持ち、ヘッジホッグ、Wnt、PI3K / mTORシグナル伝達経路の遮断を介して他の抗がん剤との相乗効果に役立つ可能性があることを示唆している。
イトラコナゾールがニーマン・ピック病C1型(NPC1)の原因タンパク質のNPC1の働きを阻害して、コレステロールの細胞内輸送を阻害し、mTORシグナル伝達と血管新生を阻害することが複数の論文で報告されています。
以下のような報告があります。
Simultaneous Targeting of NPC1 and VDAC1 by Itraconazole Leads to Synergistic Inhibition of mTOR Signaling and Angiogenesis.(イトラコナゾールによるNPC1とVDAC1の同時標的化は、mTORシグナル伝達と血管新生の相乗的阻害をもたらす)ACS Chem Biol. 2017 Jan 20;12(1):174-182.
【要旨】
抗真菌薬イトラコナゾールは、強力な抗血管新生活性を示すことが最近発見され、その後、治験中の抗がん剤として再利用されている。イトラコナゾールは、mTORシグナル伝達経路の阻害を通じてその抗血管新生活性を発揮することが示されているが、作用の分子メカニズムは不明であった。
最近、イトラコナゾールがミトコンドリアタンパク質VDAC1をターゲットとし、またmTORの上流調節因子であるAMPKの活性化のメディエーターとして作用することを明らかにした。
しかし、mTOR阻害につながることが実証されているイトラコナゾールによるコレステロール輸送阻害とVDAC1阻害作用との関連は説明できない。
この研究では、イトラコナゾールによるコレステロール輸送阻害がリソソームタンパク質NPC1の直接阻害によるものであることを明らかにした。
さらに、突然変異誘発、U18666Aとの競合、および分子ドッキングによる解析を使用して、イトラコナゾールの結合部位をNPC1のステロール感知ドメインに同定した。
最後に、AMPKの活性化とコレステロール輸送の阻害が同時に起こると、mTOR、内皮細胞の増殖、血管新生が相乗的に阻害されることを示す。
この論文はイトラコナゾールがVDAC1とNPC1を阻害する結果、mTORC1の活性を低下させて、がん細胞の増殖と血管新生を阻害するというメカニズムです。
イトラコナゾールとAMPKを活性化するメトホルミンやビタミンD3を併用すると抗腫瘍効果が高まることを示唆しています。

図:イトラコナゾールはミトコンドリアのVDAC1(Voltage-dependent anion channel-1: 電位依存性陰イオンチャネル1)を阻害し(①)、ATP産生を低下させ(②)、AMP活性化プロテインキナーゼ(AMPK)の活性を亢進し(③)、mTORC1活性を阻害する(④)。一方、イトラコナゾールはニーマン・ピック病C1型(NPC1)の原因タンパク質のNPC1を阻害し(⑤)、細胞内のコレステロール輸送を阻害する(⑥)。その結果、mTORC1の活性を阻害する(⑦)。mTORC1の活性阻害は、細胞の増殖と血管新生を阻害する(⑧)。
イタラコナゾールはニーマン・ピック病C1型(NPC1)の原因タンパク質のNPC1のステロール感知ドメイン(SSD)に結合し、NPC1によるコレステロール輸送を妨害します。その結果、リソソームでのコレステロールの蓄積と細胞内コンパートメントでのコレステロールの枯渇が誘導され、mTORC1シグナル伝達系が阻害されます。これは前述のアステミゾールと同様のメカニズムです。
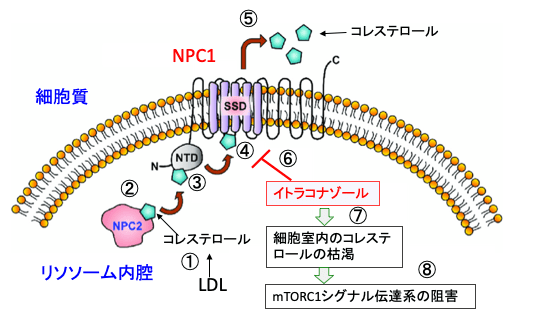
図:リソソーム内に輸送されたLDL(低密度コレステロール)はリソソームの酸性リパーゼによって加水分解されて遊離コレステロールとなる(①)。コレステロールは、リソソーム内腔のニーマンピック病C型(NPC)NPC2に結合し(②)、リソソームの内膜側にあるNPC1のN末端ドメイン(N-terminal domain:NTD)に転移する(③)。コレステロールはNPC1のN末端ドメイン(NTD)からステロール感知ドメイン(sterol-sensing domain :SSD)に転送され(④)、リソソームから細胞質に輸送される(⑤)。イトラコナゾールはNPC1のステロール感知ドメイン(SSD)に結合し、NPC1によるコレステロール輸送を妨害する(⑥)。その結果、リソソームでのコレステロールの蓄積と細胞質内のコレステロールの枯渇が誘導され(⑦)、mTORC1シグナル伝達系が阻害される(⑧)。
【VDAC-1の働きを阻害するとがん細胞の増殖がストップする】
前述の論文では、イトラコナゾールがミトコンドリアのVDAC1を阻害することが報告されています。
VDAC1(Voltage-dependent anion channel-1: 電位依存性陰イオンチャネル1)は細胞質とミトコンドリアの間の代謝産物やエネルギー(ATP)やイオンなどを通過させるゲートで、ミトコンドリア・ポリン(mitochondrial porin)とも呼ばれます。
ミトコンドリアには外膜と内膜があり、内膜の内側がミトコンドリア・マトリックス、外膜と内膜の間を膜間腔と言います。
細胞質とミトコンドリア・マトリックスの間の物質や水の行き来は、外膜と内膜に存在するチャネルやトランスポーターと呼ばれるタンパク質によって制御されています。
TCA回路で代謝されるアセチルCoAやピリピン酸やATP産生に使われるADPやリン酸は、内膜ではそれぞれ固有のトランスポーターやチャネルを使って膜を通過します。
一方、外膜では多くの物質はVDAC(Voltage-dependent anion channel:電位依存性陰イオンチャネル)を使って通過します(下図)。

図:ミトコンドリアの内膜にはアセチルCoAやピルビン酸や無機リン(Pi)やADP(アデノシン2リン酸)やATP(アデノシン3リン酸)などが通過するための特異なタンパク質(トランスポーターやチャネル)がある。ATPとADPは内膜ではANT(Adenine Nucleotide Transporter)によって通過する。ミトコンドリア外膜では、ADPやATPや無機リンや低分子物質はVDAC(Voltage-dependent anion channel:電位依存性陰イオンチャネル)を使って通過する。
このようにVDACはミトコンドリア外膜に存在する最も重要なチャネル蛋白質であり、ミトコンドリアと細胞質の間の代謝物質の輸送を制御し、細胞のエネルギー産生ひいては生存に必須の働きを行っている。(J Pharmacol Exp Ther. 342(3): 637–641. 2012年のFig.1を改変)
VDACは細胞の生存だけでなくアポトーシス(細胞死)の制御にも重要な働きをしています。すなわち、VDACはチトクロームCの通過やBCL-2ファミリー蛋白質との相互作用を制御することによってアポトーシスを制御しています。
したがって、がん治療においても、VDACは重要なターゲットになっています。
VDACには1から3まで3種類のアイソフォームがあり、分子量は30~35KDaでそれぞれ70%の類似性(homology)があります。
多くのがん細胞でVDAC-1の発現が亢進しており、VDAC-1の発現を阻害するとがん細胞の増殖が停止することが明らかになっています。
前述の論文では、イトラコナゾールがVDAC1を阻害し、ATP産生が低下すると、AMP活性化プロテインキナーゼの活性が亢進してmTORC1シグナル伝達系が抑制されるメカニズムを提唱しています。
つまり、イトラコナゾールがmTORC1シグナル伝達系を抑制するのですが、そのメカニズムとしてNPC1の阻害によるコレステロール細胞内輸送の阻害と、ミトコンドリアでのVDAC1の阻害の2つのメカニズムが関与しているという報告です。
以下のような報告もあります。
Antifungal drug itraconazole targets VDAC1 to modulate the AMPK/mTOR signaling axis in endothelial cells(抗真菌薬のイトラコナゾールはVDAC1に作用して血管内皮細胞のAMPK/mTORシグナル伝達系を制御する)Proc Natl Acad Sci U S A. 2015 Dec 29; 112(52): E7276–E7285.
イトラコナゾールが強い血管新生阻害作用を有することは多くの研究で明らかになっており、そのメカニズムとしてmTORシグナル伝達系の阻害の関与が指摘されています。しかし、イトラコナゾールのmTORシグナル伝達系阻害のメカニズムは不明です。
イトラコナゾールの血管新生阻害のメカニズムとして、イトラコナゾールが血管内皮細胞のミトコンドリアのVDAC1(電位依存性陰イオンチャネル)に結合してVDAC1の働きを阻害してATP産生を低下させ、AMP活性化プロテインキナーゼの活性を亢進してmTORシグナル伝達系を阻害して、血管新生を阻害するというメカニズムを報告しています。
つまり、VDCA1は血管新生阻害剤の重要なターゲットとなり、イトラコナゾールはVDAC1を阻害して血管新生を阻害するということです。
VDAC1遺伝子を欠損させるとmTORが阻害され、ヒト血管内皮細胞の増殖が抑制されました。VDAC1とmTORとの関連が明らかになったということです。
【がん細胞ではRAS変異によってp21活性化キナーゼ(PAK)が活性化している】
がん細胞の特徴として、細胞増殖の亢進、細胞死に対する抵抗性、周囲組織への浸潤や遠隔臓器への転移が挙げられます。
正常細胞は、増殖や細胞死や移動は厳密に制御されており、がん細胞はその制御が壊れて、無制限に増殖し、正常組織に浸潤し破壊して、増大していきます。細胞骨格はこれらのプロセスに必須の役割を担っています。
つまり、がん細胞の浸潤や転移は細胞の運動性に依存しており、細胞の運動性には細胞骨格の動的変化が必要です。
細胞の運動性を制御する細胞骨格の動的変化の背後にあるシグナル伝達経路にはRas関連の低分子量GTAアーゼ(Ras-related small GTPases)と、p21活性化キナーゼ(p21-activated kinases :PAKs)を含むエフェクタータンパク質が関与しています。
PAKキナーゼは酵母やショウジョウバエにも存在するセリン/スレオニンプロテインキナーゼのファミリーであり、哺乳類では6つのアイソフォーム(PAK1〜6)が発見されています。
それらはすべて低分子量GTPaseの RacおよびCdc42の直接の標的です。
細胞骨格の動的変化の制御における役割に加えて、PAKは細胞の生存、分裂、遺伝子転写などの様々な細胞活動を調節することが明らかになっています。
いくつかの成長因子受容体チロシンキナーゼ(インスリン、EGF、PDGF、VEGF受容体など)およびGタンパク質共役受容体からのシグナルは、PAKの活性化につながります。
これらの経路は、PI-3キナーゼ(PI3K)とグアニンヌクレオチド交換因子(GEF)の連続的な活性化を通じて低分子量GTPase のRacおよびCdc42を活性化し、PAKを活性化します。
がん細胞では、PAKの活性化は変異Rasを介して頻繁に起こっています。
Rasは最も一般的に変異しているがん遺伝子の1つであり、MAPキナーゼ経路とPI3キナーゼを活性化し、PAKキナーゼを活性化します。

図:受容体チロシンキナーゼ(インスリン受容体、EGF受容体、PDGF受容体、VEGF受容体)およびGタンパク質共役型受容体(G protein coupled receptor : GPCR)からのシグナル(①と②)は、RASおよびPI3Kを活性化し(③)、GDP/GTP交換反応を促進するGEF(guanine-nucleotide exchange factors)の作用によって(④)、Rac/Cdc42はGTP結合型になって活性化し(⑤)、Rac/Cdc42のエフェクターであるp21活性化キナーゼ(PAK-1)を活性化する(⑥)。活性化したPAK-1は細胞の生存と増殖を促進し、がん細胞の増殖と転移を亢進する(⑦)。さらに、PAK-1はRASによるMAPキナーゼ(分裂促進因子活性化タンパク質キナーゼ)経路の活性化を増強する(⑧)。PAK-1の活性化は、RacおよびCdc42を介する以外に、GTPaseに依存しないメカニズムでも起こる。
ヒトがんの70%以上において、PAK-1の活性亢進ががん細胞の増殖に関与していると言われています。
PAK1はAktを活性化してmTORC1を活性化します。
PAK1の阻害剤としてイベルメクチンが有名です。
イベルメクチン(ivermectin)はマクロライド類に属する物質で、腸管糞線虫症や糸状虫や疥癬など多くの寄生虫に有効です。
静岡県伊東市内のゴルフ場近くで採取した土壌から大村智博士により発見された新種の放線菌「ストレプトマイセス・アベルメクチニウス(Streptomyces avermitilis)」が生産するアベルメクチンを元に創製されました。大村智博士はこの発見で2015年にノーベル生理学・医学賞を受賞しています。
イベルメクチンのPAK1阻害作用については674話で解説しています。

図:低分子量Gタンパク質のRASは細胞外のさまざまな刺激(チロシンキナーゼ受容体やサイトカイン受容体やカルシウムチャネルなど)を受けて細胞増殖や生存を促進するRAFキナーゼ(Raf-1)やPI-3キナーゼ(PI3K)など多数のシグナル伝達系を活性化する(①)。p21活性化キナーゼ(PAK-1)はRacおよびCdc42のような低分子量GTPaseによって活性化される(②)。PAK-1はRasシグナル伝達系の主要な経路であるRaf-1/MEK1/ERK経路(③)とPI3K/AKT経路(④)を活性化する。さらに、PAK-1はβ-カテニンをリン酸化してWNT/β-カテニン経路を活性化する(⑤)。このように、PAK-1は複数のシグナル伝達系の制御に関与し、がん細胞の増殖と転移を促進する(⑥)。寄生虫治療薬のイベルメクチン(Ivermectin)はPAK-1とWNT/β-カテニン経路を阻害する作用によって、抗腫瘍作用を発揮する(⑦)。
以上から、メトホルミン、ビタミンD3、オーラノフィン、ラパマイシン、イトラコナゾールを併用すると、mTORC1活性を強力に抑制し、抗腫瘍活性を増強できると思います(トップの図)。
| « 735) がんにな... | 737)ミトコン... » |