がんの予防や治療における漢方治療の存在意義を考察しています。がん治療に役立つ情報も紹介しています。
「漢方がん治療」を考える
カレンダー
| 2025年1月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | |||||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| 12 | 13 | 14 | 15 | 16 | 17 | 18 | ||
| 19 | 20 | 21 | 22 | 23 | 24 | 25 | ||
| 26 | 27 | 28 | 29 | 30 | 31 | |||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
710)がん幹細胞の抗酸化システムをターゲットにしたがん治療
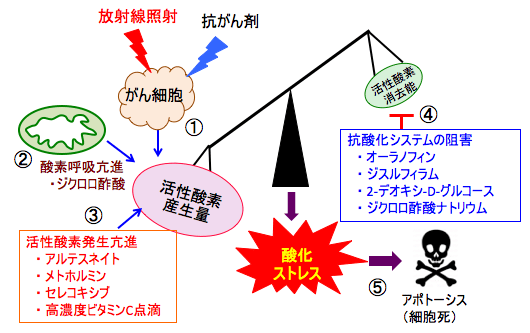
図:放射線と抗がん剤治療は活性酸素の産生を高めて細胞を死滅させる(①)。ミトコンドリアでの酸素呼吸(酸化的リン酸化)を亢進するジクロロ酢酸ナトリウム(②)、細胞内で活性酸素の産生を高める薬剤(アルテスネイト、メトホルミン、セレコキシブ、高濃度ビタミンC点滴)も活性酸素の産生を増やす(③)。活性酸素の産生量が増えると、活性酸素を消去する抗酸化物質や抗酸化酵素による抗酸化力を高めて酸化還元バランスを維持しようとする。オーラノフィン、ジスルフィラム、2-デオキシ-D-グルコース、ジクロロ酢酸ナトリウムは抗酸化システムを阻害あるいは抑制する(④)。がん細胞内の活性酸素の産生量を増やし、同時に活性酸素消去能(抗酸化力)を阻害すると、酸化還元バランスが破綻して強い酸化ストレスを引き起こし、がん細胞を死滅できる(⑤)。
710)がん幹細胞の抗酸化システムをターゲットにしたがん治療
【がん細胞は細胞内の抗酸化システムを利用して放射線照射や抗がん剤に抵抗性になる】
細胞には、活性酸素や有毒物質による害から細胞自身を守る手段や仕組みが備わっています。
例えば、細胞内で活性酸素の発生量が増えると、細胞は活性酸素を消去する酵素(スーパーオキシド・ディスムターゼ、カタラーゼ、グルタチオン・ペルオキシダーゼなど)の発現や活性を高めたり、フリーラジカルを消去するグルタチオンやチオレドキシンなどの抗酸化物質の合成を高めたりして、活性酸素の害(酸化ストレス)を軽減しようとします(下図)。
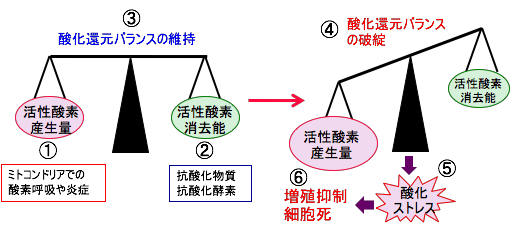
図:細胞内ではミトコンドリアで酸素を使ってATP産生を行うときに活性酸素が発生し、炎症があると炎症細胞から活性酸素が発生する(①)。このようにして産生された活性酸素は細胞に酸化傷害を引き起こすが、細胞内には活性酸素を消去する抗酸化物質や抗酸化酵素による抗酸化力が存在する(②)。活性酸素種の量と抗酸化力の差が酸化ストレスとなるが、細胞内には酸化ストレスの増大に応じて、抗酸化酵素の発現や活性を亢進することによって抗酸化力を高めるメカニズムが存在し、酸化還元のバランスを維持することによって酸化傷害の発生を防いでいる(③)。しかし、細胞内の活性酸素の産生量が増えたり、活性酸素消去能(抗酸化力)が低下すると、酸化還元バランスが破綻して(④)、酸化ストレスとなる(⑤)。酸化ストレスが亢進するとがん細胞の増殖が抑制され、細胞死が誘導される(⑥)。
放射線治療も抗がん剤治療も活性酸素の産生を高め、細胞を死滅します。
しかし、がん細胞は細胞に備わった抗酸化システムを利用して酸化ストレスを軽減し、酸化還元バランスを維持し、細胞死から免れようとします。

図:放射線や抗がん剤は、活性酸素の産生を高め(①)、細胞の酸化傷害を引き起こして、細胞増殖を抑制し、細胞死を誘導する(②)。がん細胞は、スーパーオキシド・ディスムターゼ(SOD)やカタラーゼやグルタチオン・ペルオキシダーゼなどの活性酸素消去酵素やグルタチオンやチオレドキシンなどの抗酸化物質の産生を高めて、活性酸素による害(酸化ストレス)を軽減している(③)。この抗酸化システムの亢進によって、がん細胞は放射線や抗がん剤に抵抗性になる。
そこで、ミトコンドリアでの酸素呼吸(酸化的リン酸化)を亢進したり(ジクロロ酢酸ナトリウム)、細胞内で活性酸素の産生を高める薬剤(アルテスネイト、メトホルミン、セレコキシブ、高濃度ビタミンC点滴など)を使って細胞内の活性酸素の産生量を高め、同時に、活性酸素を消去する細胞内の抗酸化システム(抗酸化力)抑制すると、細胞内の酸化ストレスが高度に亢進し、酸化傷害によってがん細胞を死滅できます。これが「がんの酸化治療」になります。(トップの図)
【がん幹細胞は抗酸化力を高めて治療抵抗性になっている】
がん組織の中にがん幹細胞 (cancer stem cells)と呼べるような細胞が存在して、通常のがん細胞を供給しながらがん組織を構成していることが明らかになっています。
従来は、がん組織に存在する全てのがん細胞が無限の自己複製能(分裂能)を有し、がん組織を形成する能力を獲得していると考えられてきました。
しかし最近の考え方は、無限の分裂能を有しがん組織を形成できるのはがん幹細胞だけであり、大部分のがん細胞は、限定された分裂能を有するか、あるいはすでに分裂能を失っていると考えられています。
また、がん幹細胞は抗がん剤や放射線治療に抵抗性であり、抗がん剤や放射線治療によって腫瘍が縮小しても、死滅しているのは分化したがん細胞だけで、がん幹細胞は生き残ることが多いことが指摘されています。
治療によって大部分のがん細胞を除いても、ごく少数のがん幹細胞が生き残っていれば再発が起こりうることになり、これが、抗がん剤治療後にしばしば再発が起きる理由だと考えられています。
現行の抗がん剤治療のほとんどは、分化したがん細胞を標的として開発されており、がん幹細胞に対してはあまり効果が無い可能性が指摘されています。
抗がん剤治療によって腫瘍が縮小しても、多くは一時的な縮小であって、がん幹細胞が生き残っているかぎり、いずれ再増殖してきます。
臨床的な奏功率(腫瘍の縮小率)が生存期間の延長に必ずしも結びつかないのは、通常の抗がん剤治療ではがん幹細胞が治療に抵抗して生き残るからだと言えます。
成熟したがん細胞が限定した分裂能しか有しないのであれば、これらは放置しておいても自然に死滅することになります。しかし、がん幹細胞が生き残っていれば、がん組織は増大し、転移も広がることになります。
つまり、がん幹細胞を効率的に死滅できないとがんは治せないということになります(下図)。
図:がん組織にはがん幹細胞 (cancer stem cells)と呼ばれる細胞が存在して、通常のがん細胞(成熟がん細胞)を供給しながらがん組織を構成している。がん幹細胞は自己複製を行う一方、不均等分裂により一部が自己複製のサイクルから逸脱して分化し通常のがん細胞となっている(①)。成熟がん細胞は抗がん剤や放射線で死滅しやすいが、がん幹細胞は死滅しにくいので抗がん剤治療や放射線治療で生き残る(②)。がん幹細胞は腫瘍形成能を持つので、生き残ったがん幹細胞が増殖して再発や再燃が起こる(③)。がん幹細胞がアポトーシス抵抗性になっているメカニズムを阻害すれば抗がん剤や放射線治療の効果を高めることができる。
がん幹細胞が分化したがん細胞よりも抗がん剤や放射線治療に対する感受性が低い理由は数多くあります。
例えば、がん幹細胞は抗がん剤の排出能力や解毒能力が高いことが指摘されています。つまり、細胞内の薬剤を排出するABC(ATP-binding cassette) transporterが高発現しているために抗がん剤が効きにくいことや、活性酸素などのフリーラジカルを消去する活性(グルタチオンやNrf2活性など)が高いために抗がん剤や放射線治療が効きにくいことが報告されています。
さらに、がん幹細胞はダメージを受けたDNAを修復する能力が高くなっているので、抗がん剤や放射線で遺伝子がダメージを受けても簡単には死ににくい性質を持っています。
がん幹細胞では、分化したがん細胞よりも、活性酸素種の量が少ないことが示されています。
通常のがん細胞よりがん幹細胞は、グルタチオンやチオレドキシンのような強力な抗酸化物質の量が多いので、その結果として細胞内の活性酸素種のレベルが低下していると考えられています。
がん幹細胞は細胞内の抗酸化物質の量が多く、そのため活性酸素種によって誘導されるアポトーシスに抵抗性を持つことになると考えられています。
がん幹細胞における抗がん剤や放射線治療に対する感受性を低下させているメカニズムを阻止すると、がん幹細胞が死滅し、がん組織を消滅できます。(下図)

図:がん組織は成熟がん細胞(①)と少数のがん幹細胞(②)から構成される。抗がん剤や放射線治療で成熟がん細胞は死にやすいが、がん幹細胞は死ににくい性質を持つので生き残る。生き残ったがん幹細胞から成熟がん細胞が産生されるので、治療後に再燃や再発が起こる(③)。薬剤の排出力や分解力の阻害、抗酸化力やDNA修復力の抑制などによってがん幹細胞の抗がん剤や放射線に対する抵抗性を阻止して(④)、がん幹細胞を死滅できればがん組織を消滅できる(⑤)。
【オーラノフィンとセレコキシブの併用による酸化ストレスの増強】
すでに使用されている薬の新規な効能を見つけて、別の疾患の治療薬にする医薬品再開発(再利用)が注目されています。
医薬品再開発は「ドラッグ・リポジショニング(Drug Repositioning)」あるいは「ドラッグ・リパーポジング(Drug Repurposing)」の日本語訳です。
「Repositioning」や「Repurposing」というのは、位置や立場(position)や目的や意図(purpose)を新たにする(re-)という意味です。医薬品の「再開発」や「再利用」という意味です。
がん幹細胞の酸化ストレスを高めてがん組織の消滅を目的とした研究においても、既存の医薬品の再利用が注目されています。以下のような報告があります。
Repurposing drugs as pro-oxidant redox modifiers to eliminate cancer stem cells and improve the treatment of advanced stage cancers.(がん幹細胞を排除し、進行期がんの治療を改善するための酸化促進性レドックス修飾剤としての医薬品の再利用)Med Res Rev. 2019 Nov;39(6):2397-2426.
【要旨】
過去10年間で、3つの大きな進歩ががんの治療効果の改善に貢献した。一つは免疫療法の進歩である。2つ目は発がんにおける分子、生化学、および細胞メカニズムの解明により、新たな薬物標的が発見されたことである。さらに3つ目は、早期発見のための信頼できるバイオマーカーの特定により、病気の早期治療が促進された。
上記の治療法の組み合わせによりがん患者の生存率が確実に改善されたにもかかわらず、単一の万能のがん治療法はまだ見つかっていない。
したがって、薬剤耐性の発生が頻繁に発生し、より悪性度の高いがん細胞の発生を促進するデメリットがあるにもかかわらず、化学療法はがん治療のための重要な治療法のままである。
ここでの焦点は、酸化ストレスを促進することによって、化学療法に対する耐性の発現を克服し、特に転移がんに対する化学療法の効き目を強化するために既存の医薬品の再利用を探ることである。
米国食品医薬品局が承認した医薬品の中で再利用に適した優れた例は、強力で特異的なチオレドキシン還元酵素阻害剤のオーラノフィン(auranofin)と非ステロイド性抗炎症薬であるセレコキシブ(celecoxib)である。
最近、この2つの薬物は、主にミトコンドリアの活性酸素種の産生を促進することにより、転移性がん細胞およびがん幹細胞を選択的に標的として、死滅することが示された。
このように進行期の転移がん細胞およびがん幹細胞の細胞内酸化還元システムをターゲットにすると、酸化ストレスを亢進し、アポトーシス(プログラムされた細胞死)の内因性経路が活性化される。
がん細胞の酸化還元システムを標的としたオーラノフィンやセレコキシブなどの医薬品の再利用によって、化学療法の奏功率と生存率の向上が期待できる。
つまり、リュウマチ治療薬として使用されているオーラノフィンと抗炎症剤のセレコキシブを組み合わせると、がん細胞の酸化ストレスが亢進して、抗がん剤治療の効き目を高めることができるということです。
がん細胞の酸化還元(レドックス)システムをターゲットにするとがん治療の効果を高めることができ、その目的にオーラノフィンとセレコキシブの組み合わせが有効という話です。
セレコキシブは関節リュウマチの治療にも使用され、オーラノフィンとセレコキシブの併用も行われています。
同様な研究は他のグループからも報告されています。以下のような論文があります。
Synergy Between Auranofin and Celecoxib Against Colon Cancer In Vitro and In Vivo Through a Novel Redox-Mediated Mechanism.( in vitroおよびin vivoでの新規レドックス媒介メカニズムによる結腸がんに対するオーラノフィンとセレコキシブの相乗効果) Cancers (Basel). 2019 Jul 3;11(7):931.
【要旨】
関節リウマチの治療のための米国食品医薬品局(FDA)承認薬であるオーラノフィンが、さまざまな実験モデルで選択的な抗がん活性を持っていることが最近の多くの研究で示されている。しかし、臨床試験の結果は、オーラノフィンは単剤として使用した場合の抗がん作用は弱いことが明らかになっている。
この研究では、オーラノフィンの抗がん活性を強化する化合物を見つけるために、FDA承認薬物ライブラリーのスクリーニングを実施し、抗炎症薬のセレコキシブがオーラノフィンの抗がん活性をin vitroおよびin vivoで強力に強化するという予想外の結果を得た。
作用機序的には、オーラノフィンとセレコキシブの組み合わせは、活性酸素の産生を増やして酸化ストレスを亢進し、活性酸素種を介したヘキソキナーゼの阻害とミトコンドリアの酸化還元恒常性の障害を引き起こして、ATP生成の大幅な減少を引き起こした。
セレコキシブによって誘導された活性酸素種産生と、オーラノフィンによるチオレドキシン還元酵素の阻害作用は、チオレドキシン還元酵素を酸化状態へシフトし、MTCO2の分解と電子輸送鎖の機能不全を引き起こす。
私たちの研究は、in vivoでがん細胞を効果的に除去する新しい薬物の組み合わせを特定した。 オーラノフィンとセレコキシブは現在臨床で使用されているFDA承認薬であるため、この研究の結果をがん治療に臨床応用することは可能である。
オーラノフィンは関節リュウマチの治療薬として古くから臨床で使用されています。前述のように、オーラノフィンはチオレドキシン還元酵素を阻害して酸化ストレスを高め、抗がん作用を発揮することが知られています。
培養細胞や動物を使った研究で抗がん作用が明らかになり、白血病、肺がん、卵巣がんなどで臨床試験が実施されています。しかし、その結果はあまり良くないようです。つまり、オーラノフィン単独での抗腫瘍効果は弱いことが明らかになっているのです。
そこで、オーラノフィンの抗腫瘍効果を高める医薬品がないかを探索する目的で、FDAが認可している1280種類の医薬品をスクリーニングしています。すでにFDAが認可している薬なので、もしオーラノフィンの抗腫瘍効果を高める効果があれば、すぐに臨床で使用できるメリットがあるわけです。
そのスクリーニングの結果、シクロオキシゲナーゼ-2の選択的阻害剤のセレコキシブがオーラノフィンの抗腫瘍効果を増強することが明らかになったのです。
この2つの組み合わせは、がん細胞のミトコンドリアの活性酸素の産生を高めて、解糖系酵素のヘキソキナーゼの活性を不活性化することが示されています。さらに、ミトコンドリアの呼吸酵素の働きを阻害してATPの産生を大きく阻害して、細胞を破綻させることを示しています。
また、セレコキシブの活性酸素産生亢進作用はCOX-2に依存しないことが示されています。すなわち、COX-2の遺伝子をノックダウンしたがん細胞でも同様な作用が見られたためです。
セレコキシブはCOX-2阻害作用以外に、この作用とは関係ないメカニズムで抗がん作用を発揮することが多く報告されています。
オーラノフィンとセレコキシブに、さらに、ジスルフィラムやジクロロ酢酸や2-デオキシ-D-グルコースやメトホルミンなどを組み合わせると、がん細胞の酸化ストレスを高めて死滅できます(下図)。

図:がん細胞はグルコーストランスポーター1(GLUT1)からのグルコースの細胞内取り込みが増えている。細胞内に取り込まれたグルコースはヘキソキナーゼでグルコース6-リン酸に変換され(①)、解糖系でピルビン酸まで代謝されたあと、乳酸脱水素酵素(LDH)で乳酸になる(②)。この解糖系では1分子のグルコースから2分子のATPが産生される(③)。ミトコンドリアに入ったピルビン酸はピルビン酸脱水素酵素(PDH)でアセチルCoAに変換され(④)、TCA回路と呼吸鎖(電子伝達系)による酸化的リン酸化によって(⑤)、大量のATP(グルコース1分子当たり約30分子のATP)が産生される(⑥)。オーラノフィンとセレコキシブを併用すると細胞内の活性酸素種(ROS)の産生が亢進し、酸化傷害によってヘキソキナーゼの活性が阻害される(⑦)。同様にミトコンドリアでも活性酸素種の産生が亢進し、呼吸鎖の酵素がダメージを受けてATP産生が阻害される(⑧)。ジスルフィラム、ジクロロ酢酸ナトリウム、2-デオキシ-D-グルコース、アルテスネイト、高濃度ビタミンC点滴、メトホルミンなども多彩なメカニズムで活性酸素種(ROS)の産生を高める(⑨)。これらの治療法を組み合わせて、がん細胞内の活性酸素種の産生を増やして酸化ストレスを亢進すると細胞死を誘導できる(⑩)。
オーラノフィンはがん幹細胞の排除に有効であることが注目されています。以下のような報告があります。
Elimination of Stem-Like Cancer Cell Side-Population by Auranofin Through Modulation of ROS and Glycolysis(活性酸素種と解糖系の調節によるオーラノフィンによる幹細胞様がん細胞のサイドポピュレーションの排除)Cell Death Dis. 2018 Jan 24;9(2):89.
【要旨】
がんのサイド・ポピュレーション(side-population)は、薬物の輸出に関与するATP結合カセットトランスポーターABCG2の発現が高いため、薬物耐性に重要な役割を果たす幹様がん細胞の亜集団を表す。
関節リウマチの治療に使用される金錯体の医薬品であるオーラノフィンは、腫瘍の増殖を抑制する効果が報告されている。
しかし、オーラノフィンがサイドポピュレーション細胞に影響を与えることができるかどうかは不明である。
我々の研究は、オーラノフィンがサイドポピュレーション細胞の枯渇と幹細胞マーカーのダウンレギュレーション(発現低下)を引き起こし、in vitroで腫瘍コロニーを形成する能力と、in vivoで肺がんの腫瘍発生を抑制することを示した。
これらの作用は抗酸化剤のNAC(N-アセチルシステイン)で阻害されたことから、活性酸素種がオーラノフィンによるサイドポピュレーション細胞の枯渇を媒介する重要な役割を果たしていることが示された。
さらなる研究により、オーラノフィンは解糖の阻害によりATPの減少を引き起こす可能性があることが明らかになった。
細胞ATPの枯渇はABCG2ポンプの機能を損ない、細胞内の薬物蓄積の増加につながり、したがってアドリアマイシンなどの化学療法剤の抗癌活性を高める可能性がある。 オーラノフィンとアドリアマイシンの相乗効果は、in vitroとin vivoの両方で実証された。
活性酸素種の増加と解糖の阻害は、幹細胞様のがん細胞を排除するための新しい戦略となる。 オーラノフィンとアドリアマイシンの併用は、治療効果を高めることが期待できる。
多くの幹細胞がHoechst 33342というDNA蛍光色素に対して高い排出能をもつことが報告されており、Side Population (SP) 細胞とよばれています。
つまり、Side Population細胞というのは、DNA結合色素であるHoechst 33342に対して低染色性を呈する細胞です。このSP細胞は高い幹細胞活性を持つことが知られています。
オーラノフィンはこのSP細胞のABCG2ポンプの機能を低下させてがん幹細胞活性を抑制するという報告です。
【がん幹細胞ではペントースリン酸経路が亢進している】
がん細胞は正常細胞に比べて細胞増殖が亢進しています。
したがって、がん細胞を死滅させる手段としては、DNAの合成や複製の過程、細胞分裂のメカニズム(微小管の働きなど)がターゲットになります。
すなわち、細胞傷害性の抗がん剤の多くは、細胞分裂の過程を阻害して、がん細胞の増殖を抑え、細胞死を誘導することを目標にしています。
しかし、正常細胞でも、骨髄細胞や消化管粘膜上皮細胞や免疫組織(リンパ球)や毛根細胞も盛んに細胞分裂を行っているので、細胞増殖を阻害する抗がん剤は、骨髄抑制(白血球減少、血小板減少、貧血)や消化管障害(食欲低下、吐き気、嘔吐、便通障害など)や免疫力低下(リンパ球減少)や脱毛などの副作用が出てきます。
ある特定の分子を標的として、その機能を制御することにより治療する分子標的薬も多く開発されています。がんの分子標的薬の多くは、細胞の増殖や生存のシグナル伝達系のタンパク質がターゲットになっています。
しかし、複数の経路が相互にクロストークしながらネットワークを形成しているので、シグナル伝達経路の一部を阻害しても、迂回経路の存在によって、がん細胞の増殖を十分に阻害することに限界があり、したがってその効果は限定的です。
この他のがん治療のターゲットとしては、「エネルギー産生と物質合成」や「抗酸化システム」があります。
前者に関しては、解糖系やペントース・リン酸経路やミトコンドリアがターゲットになります。
後者に関しては、グルタチオンやチオレドキシンなどがターゲットになります。
解糖系とペントース・リン酸経路とミトコンドリアと抗酸化システム(グルタチオンやチオレドキシン)を同時にターゲットにしたがん治療は、正常細胞へのダメージは少なく、がん細胞に選択的に作用して、増殖抑制と細胞死誘導の効果が期待できます。
がん幹細胞ではペントース・リン酸経路と抗酸化システムが亢進している事が明らかになっています。以下のような論文があります。
Cancer Cells Change Their Glucose Metabolism to Overcome Increased ROS: One Step From Cancer Cell to Cancer Stem Cell?(がん細胞は、増加した活性酸素種を克服するために、グルコース代謝を変更する:がん細胞からがん幹細胞への1つのステップ?)Biomed Pharmacother. 2019 Apr;112:108690.
【要旨】
がん細胞は、ATP枯渇による低エネルギー状態や活性酸素種の過剰状態に、代謝およびエネルギー産生ネットワークを変更することにより適応できる。
エネルギー枯渇や酸化ストレスは細胞の生存や薬剤耐性の獲得にも役割を果たす可能性がある。
がん細胞は一般に解糖の増加を特徴としている。さらに、 がん幹細胞は、ペントースリン酸経路の代謝の亢進を示す。
最近の研究から、酸化ストレスが亢進したとき、がん細胞は最初に解糖を増加させ、酸化ストレスがさらに持続すると、代謝バランスが解糖からペントースリン酸経路に移行する。
持続的な酸化ストレス影響下において、このような代謝の変更は、がん細胞の幹細胞へのシフトの兆候である可能性がある。
がん幹細胞では、ペントースリン酸経路が亢進しているので、ペントースリン酸経路の阻害はがん幹細胞をターゲットした治療法の一つになります。そこで、ペントースリン酸経路について理解することががん治療の理解に役立ちます。
【ペントースリン酸経路は核酸の原料と還元剤のNADPHを産生する】
ペントースリン酸経路とは、解糖系の中間体のグルコース6リン酸から分岐し、同じく解糖系の中間体 グリセルアルデヒド3リン酸に戻る経路(回路)です。解糖系と同様に細胞質に存在する経路で、補酵素の一つであるNADPHを産生し、核酸の原料となるリボース5リン酸などの5単糖 (ペントース) を産生します(下図)。

図:解糖系は1分子のグルコースが2分子のピルビン酸に分解される過程で2分子のATPが産生される(①)。グルコース6リン酸から派生するペントースリン酸経路では、還元剤のNADPHが2分子産生され、グルタチオン還元や脂肪酸合成など還元力を必要とする生合成反応に使われる(②)。さらに、核酸合成の材料になるリボース5リン酸が産生される(③)。がん細胞ではグルコースの取込みが増え、解糖系とペントースリン酸経路が亢進して、細胞分裂のためのエネルギー(ATP)と物質合成(核酸、脂肪酸、NADPHなど)が亢進している。
NADPH(ニコチンアミドアデニンジヌクレオチドリン酸、nicotinamide adenine dinucleotide phosphate)は還元剤です。脂肪酸やステロイドの合成、抗酸化物質のグルタチオンやチオレドキシンの還元剤として使用されます。
解糖系はATPを産生します。ペントースリン酸経路はATP産生には関与せず、核酸の原料や還元剤(NADPH)の産生を行っています。
細胞が増殖するにはエネルギー(ATP)だけでなく、核酸や脂肪酸などの物質合成や、酸化ストレスを軽減する還元剤の需要も増えます。したがって、がん細胞では、解糖系とペントースリン酸経路が亢進しています。
生存するだけであれば、エネルギー産生ができればよいので、解糖系だけで十分かもしれません。
しかし、細胞分裂するためには、核酸や脂肪酸やタンパク質などの物質を合成する必要があります。
がん幹細胞は細胞を増やす細胞であるから、物質合成を増やす必要があり、その目的のためにペントースリン酸経路が亢進していると言えます。
【還元型グルタチオンと酸化型グルタチオン】
グルタチオン(Glutathione)というのは、グルタミン酸とシステインとグリシンの3つのアミノ酸が結合したトリペプチドです。
γ-グルタミルシステイン合成酵素によってグルタミン酸とシステインが結合してγ-グルタミルシステインを合成します。引き続いてグルタチオン合成酵素によってγ-グルタミルシステインにグリシンが結合してグルタチオンが合成されます。この合成にはATPが必要です。
つまり、グルタミン酸やシステインやグリシンが不足したり、ATPが十分に産生できなかったり、γ-グルタミルシステイン合成酵素やグルタチオン合成酵素の活性が阻害されれば、グルタチオンの濃度は低下して、酸化ストレスに対する抵抗力が低下することになります。

図:グルタチオンは3つのアミノ酸(グルタミン酸、システイン、グリシン)がATPを使って結合して合成される。
グルタチオンは細胞内に0.5〜10mMという非常に高濃度で存在します。チオール基(SH基)を持ち、この水素が電子を供与することによって活性酸素やフリーラジカルを消去します。
還元型のグルタチオンはGSH(Glutathione-SH)と表記され、GSHが活性酸素などで酸化されると酸化型グルタチオンGSSG(Glutathione-S-S-Glutathione)になります。
つまり、酸化型は、二分子の還元型グルタチオンがジスルフィド結合(2個のイオウ原子が繋がった状態)によってつながった分子です。
細胞内で発生した活性酸素やフリーラジカルに電子を与えて酸化型になったグルタチオンを還元型に戻す酵素がグルタチオンレダクターゼ(グルタチオン還元酵素)で、このときNADPH(ニコチンアミドアデニンジヌクレオチドリン酸、nicotinamide adenine dinucleotide phosphate)から水素をもらいます。このNADPHはペントース・リン酸経路で産生されます。

図:還元型グルタチオンは活性酸素(スーパーオキサイド、過酸化水素など)などと反応して酸化され、2量体化した酸化型グルタチオン(GSSG)に変化するが、グルタチオン還元酵素がNADPHからの電子をGSSGに転移して、GSH(還元型グルタチオン)に再生される。
がん細胞は還元型グルタチオン(GSH)の産生を促進することで、酸化ストレス抵抗性を高め、増殖や転移や治療抵抗性を高めていることが知られています。
NADPHはペントース・リン酸経路で産生されます。つまり、がん細胞のグルコース取り込みや解糖系やペントース・リン酸経路を阻害するケトン食や2−デオキシ-D-グルコースやジクロロ酢酸はNHDPHの供給を減らすことによって、グルタチオンの合成を低下させ、酸化ストレスに対する抵抗性を減弱させることができます。
ATP産生低下はグルタチオン合成をさらに低下させます。また、NADPHの産生抑制は脂肪酸合成を抑制して細胞増殖を低下します。
【ジクロロ酢酸ナトリウムと2-デオキシグルコースはペントースリン酸経路を阻害する】
がん治療において、還元剤と核酸の原料を供給するペントースリン酸経路の阻害は重要な治療法になります。
ピルビン酸脱水素酵素キナーゼを阻害してミトコンドリアを活性化するジクロロ酢酸ナトリウムがペントースリン酸経路を阻害することが報告されています。以下のような論文があります。
Inhibition of the pentose phosphate pathway by dichloroacetate unravels a missing link between aerobic glycolysis and cancer cell proliferation(ジクロロ酢酸によるペントースリン酸経路の阻害は、好気性解糖とがん細胞増殖との間の失われた関連を明らかにする)Oncotarget. 2016 Jan 19; 7(3): 2910–2920.
【要旨】
がん細胞は酸素の存在下でも解糖によるグルコースの発酵を行っており、これはワールブルグ(Warburg)効果と呼ばれている。このワールブルグ効果は、がんの治療法の開発において魅力的なターゲットになっているがん細胞に共通の特徴である。
本研究は、がん細胞における代謝、エネルギー貯蔵および増殖速度の間の関係を分析することを目的とした。6つのがん細胞株において、DNA合成量によって評価した細胞増殖能は、解糖の効率と相関することを見出した。
解糖と増殖の関係をさらに調べるために、ペントースリン酸経路の薬理学的阻害を使用した。
我々は、ペントースリン酸経路の活性の低下ががん細胞の増殖を減少させ、その作用はワールブルグ効果の代謝が強いがん細胞ほど大きな影響を及ぼすことを実証した。
ペントースリン酸経路の最初の律速酵素であるグルコース-6-リン酸デヒドロゲナーゼに対するsiRNAを用いて、がん細胞の増殖を維持する上でのペントースリン酸経路の重要な役割が確認された。
さらに、ジクロロ酢酸が、がん細胞の解糖系優位の代謝からミトコンドリアでの酸化的リン酸化を亢進するように代謝を変換させ、それに応じて増殖能が減少することを見出した。
ジクロロ酢酸がペントースリン酸経路の活性を低下させたことを実証することにより、ジクロロ酢酸ががん細胞の増殖を制御する新しいメカニズムを提供する。
正常細胞では解糖と酸化的リン酸化が連動して働き、ATPを産生しています。
がん細胞では解糖と酸化的リン酸化が連動していません。解糖の最終産物のピルビン酸は乳酸に変換され、ミトコンドリアでの酸化的リン酸化は抑制されています。
増殖する細胞にとっては、エネルギー産生と物質合成を両立させるためにはグルコースの取込みを亢進し、解糖系とペントースリン酸経路を亢進する必要があります。
ジクロロ酢酸はミトコンドリアの酸化的リン酸化を促進し、その結果、解糖系とペントースリン酸経路を抑制する結果になります。
つまりジクロロ酢酸と2−DGで解糖系とペントースリン酸経路を抑制すると、ATP産生と物質合成を低下させて、がん細胞の増殖を抑制できます。(下図)

図:2−デオキシ-D-グルコース(2-DG)はヘキソキナーゼ(HK)で2-DG-6リン酸(2-DG-6-PO4)に変換される(①)。2-DG-6リン酸はヘキソキナーゼ(HK)とホスホグルコースイソメラーゼ(PGI)を阻害して解糖系とペントースリン酸経路を阻害する(②)。ジクロロ酢酸ナトリウムは、ピルビン酸脱水素酵素を活性化してピルビン酸からアセチルCoAの変換を亢進してミトコンドリアでの代謝を亢進し、解糖系とペントースリン酸経路を阻害する(③)。解糖系とペントースリン酸経路の阻害は、エネルギー産生や物質合成や抗酸化システムを阻害してがん細胞の増殖を抑制し、細胞死を誘導する。
【解糖系とペントースリン酸経路と抗酸化システムを同時に阻害するとがん細胞を選択的に死滅する】
解糖とペントースリン酸回路と抗酸化システム(グルタチオンやチオレドキシン代謝やNrf2活性など)を同時に阻害すると、がん細胞に選択的に酸化ストレスを高めて増殖を阻害することが報告されています。以下のような報告があります。
Combined inhibition of glycolysis, the pentose cycle, and thioredoxin metabolism selectively increases cytotoxicity and oxidative stress in human breast and prostate cancer(解糖系とペントースリン酸回路とチオレドキシン代謝の同時阻害は,ヒトの乳がんと前立腺がんにおいて細胞毒性と酸化ストレスを選択的に亢進する)Redox Biol. 2015 Apr; 4: 127–135.
【要旨】
ヒト前立腺がん細胞株(PC-3とDU145)とヒト乳がん細胞株(MDA-MB231)を使った実験系において、培養細胞の解糖系を2-デオキシ-d-グルコース(2DG; 20mM, 24-48 hr)で阻害し、同時にペントースリン酸回路をデヒドロエピアンドロステロン(DHEA, 300μM, 24-48 hr)で阻害すると、チオール基を介した酸化ストレスの亢進によって細胞死が誘導できる。
驚くべきことに、2DG+DHEAを培養液に添加するときに、細胞内のグルタチオン量を90%枯渇させる濃度のグルタチオン合成の阻害薬(l-buthionine sulfoximine; BSO, 1mM)を同時に添加して48時間経過しても、2DG+DHEAでみられた細胞死のレベルを増強することはなかった。
これとは対象的に、チオレドキシン還元酵素の活性を阻害するオーラノフィン(Auranofin; 1μM)を2DG+DHEA あるいはDHEAのみを添加した培養液に添加すると、3つの細胞株全てにおいて、24時間後には細胞死は顕著に増加した。
さらに、DHEA+オーラノフィンの組合せによって増加した細胞死誘導は、チオールの抗酸化剤であるN-アセチルシステイン(NAC, 20mM)の添加によってほぼ完全に阻止された。
PC-3細胞を用いた解析で、DHEA+オーラノフィンの併用投与でチオレドキシン-1の酸化が促進され、N-アセチルシステインによって阻害される事が示された。
重要な点は、正常なヒト乳腺上皮細胞は2DG+DHEA+オーラノフィンの組合せに対して、がん細胞のような細胞死が起こらなかった。
これらの結果は、解糖系とペントースリン酸回路とチオレドキシン代謝を同時に阻害する方法は、がん細胞に選択的に酸化ストレスを誘導して死滅させる有効な手段となる可能性を示唆している。
DHEA(デヒドロエピアンドロステロン)はペントース・リン酸回路でNADPHを産生するグルコース-6-リン酸デヒドロゲナーゼ(G6PD)を阻害する作用があります。
この実験は培養細胞を使ったin vitroの実験であるため、この結果をそのまま生体内の効果に結びつけることはできません。
その理由は、この実験で使用されている20mMの2-デオキシグルコース(2-DG)と300μMのDHEA(デヒドロエピアンドロステロン)は生体内では達成できない濃度だからです。
しかし1μMのオーラノフィンの濃度は現実的な数値であり、関節リュウマチの治療に使用される服用量で生体内の細胞のチオレドキシン還元酵素を阻害することは可能です。
また、ケトン食や2-DGやジクロロ酢酸などを併用するとがん細胞に選択的にがん細胞の解糖系とペントースリン酸回路の阻害は可能です。
また、グルタチオンの産生を阻害するスルファサラジン(サラゾスルファピリジン)の併用はオーラノフィンの抗腫瘍効果を高めることができます。
抗酸化システムの阻害はがん細胞の抗がん剤感受性を高めます。以下のような報告があります。
Enhancement of carboplatin-mediated lung cancer cell killing by simultaneous disruption of glutathione and thioredoxin metabolism.(グルタチオンとチオレドキシン代謝の同時阻害による肺がん細胞に対するカプボプラチンの抗腫瘍効果の増強)Clin Cancer Res. 2011 Oct 1;17(19):6206-17.
【要旨】
目的:正常細胞に比べて、がん細胞では細胞内のヒドロペルオキシド(過酸化物)のレベルが定常的に高くなっている。そのため、がん細胞内のグルコース代謝とヒドロペルオキシド代謝が亢進している。本研究では、グルコースとヒドロペルオキシドの代謝の阻害剤が、肺がん細胞において酸化ストレスを高めることによって、抗がん剤や放射線の感受性を高めるかどうかを検討した。
実験計画:ヒト肺がん細胞株のA549およびNCI-H292を用い、カルボプラチン+電離放射線と2-デオキシ-d-グルコース(2DG)の組合せで処理した。さらに、in vitroおよびin vivoの実験系において、グルタチオン合成阻害薬のブチオニンスルホキシミン(buthionine sulfoximine)およびチオレドキシン依存性代謝の阻害薬のオーラノフィンを用いて、カルボプラチンおよび電離放射線に対する肺がん細胞の感受性を高めた。
結果:2DGをカルボプラチン+ 電離放射線と組み合わせた場合、A549細胞およびNCI-H292肺がん細胞において細胞死滅が増強され、この組み合わせはパクリタキセル+カルボプラチン+ 電離放射線より有効であった。
チオール性抗酸化剤(N-アセチルシステイン)は、2DG +カルボプラチン誘発細胞殺傷からがん細胞を保護することができた。単一の薬剤として毒性の出ない用量でのブチオニンスルホキシミンおよびオーラノフィンの同時投与は、肺がん細胞の2DG +カルボプラチンに対する感受性を増強した。
この治療の組合せは、グルタチオンとチオレドキシンの両方の酸化を増加させ、この作用はN-アセチルシステインによって阻害された。
オーラノフィン+ ブチオニンスルホキシミンで処置したマウスは、血液中の白血球または赤血球に変化を示さなかった。
マウスの移植腫瘍を用いた実験系では、カルボプラチンまたはオーラノフィン+ ブチオニンスルホキシミンで処置した動物よりも、オーラノフィン+ブチオニンスルホキシミン +カルボプラチンでの処置によってより効果的に腫瘍増殖が阻害された。
結論:これらの結果は、in vitroとin vivoの実験系において、グルタチオンおよびチオレドキシン代謝の同時阻害が、肺がん細胞の増殖を効果的に阻害し、チオール媒介性酸化ストレスを含む機構によって、抗がん剤および放射線に対する感受性を高めることができることを示している。
以上の報告から、抗がん剤治療や放射線治療に解糖系とペントースリン酸経路と抗酸化システムを阻害する治療の併用が有効であることが理解できます。
抗がん剤治療や放射線治療と併用しなくても、がん細胞に十分に酸化ストレスを高めればがん細胞の増殖を抑制し、細胞死を誘導できます。
ミトコンドリアでの酸素呼吸(酸化的リン酸化)を亢進したり(ジクロロ酢酸ナトリウム)、細胞内で活性酸素の産生を高める薬剤(高濃度ビタミンC点滴、アルテスネイト、メトホルミン、セレコキシブ)を使ってー細胞内の活性酸素の産生量を高め、同時に、活性酸素を消去する細胞内の抗酸化システム(抗酸化力)を抑制する薬剤(オーラノフィン、ジスルフィラム、2-デオキシ-D-グルコース)を併用すると、細胞内の酸化ストレスが高度に亢進し、酸化傷害によってがん細胞を死滅できます。(トップの図)
| « 709)抗がん剤... | 711)代謝をタ... » |