がんの予防や治療における漢方治療の存在意義を考察しています。がん治療に役立つ情報も紹介しています。
「漢方がん治療」を考える
カレンダー
| 2024年12月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| 8 | 9 | 10 | 11 | 12 | 13 | 14 | ||
| 15 | 16 | 17 | 18 | 19 | 20 | 21 | ||
| 22 | 23 | 24 | 25 | 26 | 27 | 28 | ||
| 29 | 30 | 31 | ||||||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
594) Hippo経路をターゲットにしたがん治療(その1):シンバスタチンとセレコキシブと肝臓がん
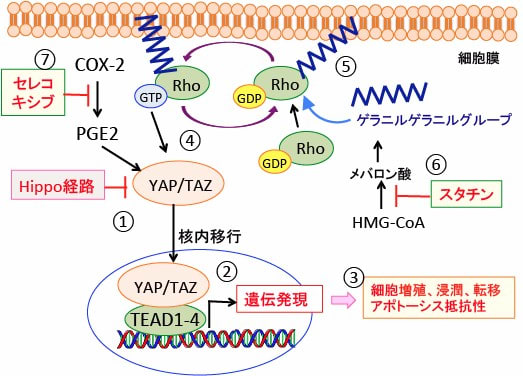
図: Hippo経路はYAP/TAZの核内移行を阻止している(①)。Hippo経路が不活性化されると、YAP/TAZは核内に蓄積し、TEADと相互作用して、YAP/TAZのターゲット遺伝子の発現を誘導する(②)。その結果、がん細胞の増殖や転移が促進される(③)。低分子量Gタンパク質の一種のRhoはYAP/TAZを活性化する(④)。RhoがGTP結合型の活性型に変換されるには、Rhoがゲラニルゲラニルグループによって細胞膜の脂質に接着していなければならない(⑤)。ゲラニルゲラニルグループは、ヒドロキシメチルグルタリル-CoA(HMG-CoA)からメバロン酸経路で産生される。HMG-CoAからメバロン酸を合成するHMG-CoA還元酵素の阻害剤であるスタチンはゲラニルゲラニルグループの産生を阻害することによってRhoの活性化を阻害する(⑥)。シクロオキシゲナーゼ-2(COX-2)はプロスタグランジンE2(PGE2)を介してYAPの発現を亢進するので、COX-2阻害剤のセレコキシブ(celecoxib)はYAPの働きを抑制する。
594) Hippo経路をターゲットにしたがん治療(その1):シンバスタチンとセレコキシブと肝臓がん
【がん研究における接触阻害(contact inhibition)の研究は長く停滞していた?】
私が病理でがんの研究を始めた40年くらい前は、がん細胞の特徴として「接触阻害(contact inhibition)」という用語を使う頻度は今より高かったように思います。接触阻害というのは、細胞がお互いに接触するまで増殖した場合に細胞の増殖が停止する現象です。
しかし、しばらくの間、がん研究領域で「接触阻害」の用語を聞く頻度が少なかったように思います。このブルグでも、「接触阻害」や「接触阻止」で検索しても何もヒットしません。つまり、がん細胞の重要な性質である「接触阻害」をいう用語を過去593回で一度も記述していないことに最近気がついて驚いた次第です。
PubMedで「contact inhibition cancer」で検索してヒットする論文は1960年代から見つかり、1974年に61件で一旦ピークになり、その後1999年までの25年間は1年間に30〜40件程度で、1983年は18件で最低になっています。2000年以降も50〜70件程度であまり増えていません。
様々な増殖因子とその受容体によるシグナル伝達系が明らかになり、増殖因子による細胞増殖の制御や、細胞死(アポトーシス)のメカニズムの解明が進んだので、機械的な接触による細胞増殖に制御に関する研究は相対的に少なくなったのかもしれません。多くの研究者が増殖因子とその受容体による増殖シグナル伝達系や細胞死のメカニズムを研究のターゲットにしていたからです。
ところが、最近は、この接触阻害のメカニズムが急速に明らかになっています。
【がん細胞は接触阻害(contact inhibition)による増殖停止ができない】
がん細胞は増殖や生存に関連する遺伝子の異常によって、増殖の制御が壊れたために「無制限に増殖する」ようになった細胞です。
正常な細胞は隣り合う細胞同士が互いに接すると、細胞増殖を停止させる接触阻害(コンタクト・インヒビション)という現象がみられますが、悪性腫瘍ではこの現象が破綻しており、増殖停止が起こらず、無制限な細胞増殖をします(下図)。
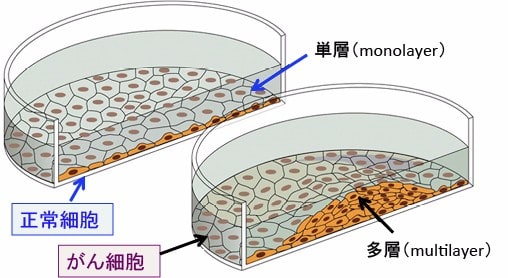
図:シャーレで細胞を培養すると、正常細胞はシャーレに細胞が一杯になったとき、細胞同士が接触阻害(contact inhibition)によって増殖を停止するので、単層(monolayer)の状態で細胞増殖は停止する。一方、がん細胞では、細胞同士の接触阻害による増殖停止の機序が起こらないので、幾重にも細胞が重なって栄養分があるかぎり増殖を続ける。
【接触阻害を制御するHippo経路】
接触阻害(コンタクトインヒビション)を制御するメカニズムの中心がHippo経路です。Hippo経路は器官のサイズを制御するシグナル経路としてショウジョウバエで発見されました。
Hippo 経路はほ乳類にも高度に保存され、器官のサイズや形の制御の他にも、がん抑制経路として、また細胞分化に重要な経路として認識されるようになり、がん治療のターゲットとして近年注目されています。
典型的なほ乳類Hippo 経路ではMST、LATS の2つのキナーゼとそれぞれアダプター分子SAV、MOB1の4つをコア構成分子とし、転写共役因子であるYAP(Yes-associated protein)やTAZ(transcriptional co-activator with PDZ-binding motif) がその下流で作動します。
刺激をうけて活性化したMST キナーゼはLATS キナーゼをリン酸化し、活性化されたLATS キナーゼは,主に細胞増殖に働くYAP/TAZ をリン酸化します。リン酸化されたYAP/TAZ は核から細胞質へ排出されるとともに、タンパク質分解を受けます。
すなわち,Hippo 経路の活性化はYAP/TAZ を負に制御し、細胞増殖抑制に働くのです。
YAPとTAZ自身には転写因子活性はなく、転写因子のTEADs(TEA domain transcription factors)に転写共役因子として作用して、転写促進作用を発揮します。
YAP/TAZは、細胞増殖や組織傷害後の組織再生を促進する遺伝子の発現を亢進します。したがって、再生医学ではYAP/TAZの活性化が治療に使われます。しかし、がん細胞では、YAP/TAZはがん細胞の増殖を促進することになり、YAP/TAZの活性化を阻害することが治療になります。

図:(左)Hippoシグナル経路がONのとき、LATS1/2がYAP/TAZをリン酸化する。リン酸化したYAP/TAZは分解するか、14-3-3と結合して細胞質内に保持される。この場合は、YAP/TAZによる遺伝子転写は阻止される。
(右)Hippoシグナル経路が不活性化されると、YAP/TAZは核内に蓄積し、TEADと相互作用して、YAP/TAZのターゲット遺伝子の発現を誘導する。(参考:Genes 2016, 7(9), 55;Targeting the Hippo Signaling Pathway for Tissue Regeneration and Cancer Therapy)
【肝臓がんではYAP/TAZの活性化が高頻度に起こっている】
正常な肝臓を切除すると、残った肝細胞が増殖し、元の大きさ(細胞数)に達すると細胞の増殖は停止します。肝臓がその大きさを知るメカニズムに接触阻害のメカニズムが関与していることは予想できますが、その具体的なメカニズムは長く不明でした。しかし、肝臓の大きさの決定にHippo経路が関与していることが明らかになっています。
マウスの肝細胞でYAPを過剰に発現させると、肝細胞や未熟胆管細胞の過形成のために肝臓が著しく腫大し、肝がんを発症します。
一方で肝細胞特異的なYAP欠損マウスでは、肝細胞や胆管細胞の減少と肝がんの発症抑制が報告されています。またYAPの活性化は、成熟肝細胞を前駆細胞に脱分化させ、1細胞レベルで自己複製能や可移植能を持つようになることも示されています。
さらにYAP/TAZの活性を抑える上流のキナーゼやアダプタータンパク質(MST1/2、SAV1、LATS1/2、MOB1A/1B)のいずれかを肝臓特異的に欠損させたマウスにおいても、肝腫大と肝がん発症促進が認められることが報告されています。
つまり、Hippo-YAP/TAZ経路の破綻(YAP/TAZの活性化)は、肝臓を腫大させ、肝臓がんの発生を促進する結果を引き起こします。
つまり、Hippo 経路は肝細胞の増殖停止や器官サイズの制御に重要であり、その破綻は肝臓幹細胞や前駆細胞への脱分化と、腫瘍化を引き起こすことになります。
したがって、Hippo-YAP/TAZ経路は肝臓がん治療の重要なターゲットになります。
臨床例での研究では、肝細胞がんと肝内胆管がんとヘパトブラストーマ(肝芽腫)において、免疫染色において細胞核内のYAPの量が増えていることが報告されています。そして、核内のYAPの量が多いほど、がん細胞の悪性度が高く、予後が不良であることが明らかになっています。
YAP発現と血清中のAFPレベルが相関するという報告もあります。さらに、YAP発現の高いがん細胞では抗がん剤耐性が強くなることが報告されています。
【シンバスタチンはTAZを抑制して肝細胞がんの増殖を抑える】
高脂血症の治療薬のスタチンがHippo-YAP/TAZ経路に作用して肝臓がんの増殖を抑えることが報告されています。以下のような報告があります。
Statin attenuates cell proliferative ability via TAZ (WWTR1) in hepatocellular carcinoma.(スタチンは、肝細胞がんにおけるTAZ(WWTR1)を介した細胞増殖能を弱める)Med Oncol. 2016 Nov;33(11):123.
熊本大学医学部の消化器外科の研究グループからの論文です。
メバロン酸経路とHippo経路には相互に関連(クロストーク)があることが報告されています。
高脂血症治療薬のスタチン(Statins)はメバロン酸経路を阻害します。そこで、この研究では、肝細胞がんに対するスタチン(Statins)の抗腫瘍効果を検討しています。
肝細胞がん細胞株を用いた実験では、4種類のスタチン(atorvastatin, pravastatin, fluvastatin, simvastatin)のうち、シンバスタチン(Simvastatin)とフルバスタチン(Fluvastatin)の2つが最も強い増殖抑制作用とアポトーシス誘導作用を示し、TAZの発現がこの2種類のスタチンで抑制されることを示しています。
さらに、治癒切除を受けた肝細胞がん患者における検討で、スタチンを服用していた患者のグループは、スタチンを服用していない患者グループより無再発生存期間が有意に長いことを報告しています。
以上から、この論文の結論は「スタチンは肝細胞がん細胞に対して増殖抑制と細胞死(アポトーシス)を誘導し、肝細胞がん患者の予後を改善する」となっています。
つまり、肝臓がん患者はシンバスタチン(Simvastatin)かフルバスタチン(Fluvastatin)を服用するメリットがある可能性を示唆しています。
前述のように、Hippo-YAP/TAZシグナル伝達経路において、Hippoシグナル伝達経路が破綻すると転写共役因子のYAPおよびTAZが核内に移行して、細胞増殖に関連する遺伝子の転写を亢進します。
Hippo-YAP/TAZ経路は肝臓のサイズを制御するシグナル伝達経路の中心です。肝臓がんではHippo-YAP/TAZ経路の異常が高頻度に認められています。
たとえば,YAPは肝がんの患者の約60%、TAZは肝がんの患者の約50%で高発現や核への局在が認められるという報告があります。そしてYAPやTAZの高発現や核への局在亢進が予後不良と関連していることが報告されています。
YAP/TAZの活性化に必要なGTP結合タンパク質(Gタンパク質)のイソプレニル化はメバロン酸経路で産生される物質が必要です。したがって、メバロン酸経路を阻害するスタチンはYAP/TAZの活性化を阻害することになります。
メバロン酸経路の阻害によるYAP/TAZ阻害のメカニズムを以下に解説します。
【スタチンは肝臓でのコレステロールの合成を抑制する】
コレステロールは、アセチルCoA(グルコースや脂肪酸やグルタミンなどの分解によって生成される)から生成されるメバロン酸(Mevalonic acid)を経て生合成されます。この生合成経路をメバロン酸経路と言い、この経路の律速酵素である3-ヒドロキシ-3-メチルグルタリルCoA還元酵素(3-hydroxy-3-methylglutaryl-CoA reductase ;HMG-CoA還元酵素)を阻害すると肝臓でのコレステロール生合成を抑制することができるため、多くのHMG-CoA還元酵素阻害剤が開発され高脂血症治療薬として臨床で使われています。
このようなHMG-CoA還元酵素の働きを阻害することによって血液中のコレステロ-ル値を低下させる薬(HMG-CoA還元酵素阻害剤)の総称をスタチン(Statin)といいます。

図:スタチンは肝臓においてヒドロキシメチルグルタリル-CoA(HMG-CoA)からメバロン酸に変換するHMG-CoA還元酵素を阻害することによってコレステロール合成を抑制する。
最初のスタチンであるメバスタチンは1973年に青カビの一種から発見され、それ以降、様々な種類のスタチンが開発され、高コレステロール血症の治療薬として世界各国で使用されています。近年の大規模臨床試験により、スタチンは高脂血症患者での心筋梗塞や脳血管障害の発症リスクを低下させる効果があることが明らかにされています。
スタチンのHMG-CoA還元酵素に対する親和性(affinity)は本来の基質であるHMG-CoAの1000倍以上であるため、HMG-CoA還元酵素を強力に阻害します。
【スタチンはメバロン酸経路でのイソプレノイド生成を低下してGTP結合タンパク質の活性を阻害する】
メバロン酸はコレステロールの合成に必要なだけでなく、GTP結合タンパク質(Gタンパク質)のイソプレニル化に必要な物質(geranylpyrophophateやfarnesylpyrophosphate)を作ります。(下図)

図:ヒドロキシメチルグルタリル-CoA(HMG-CoA)からメバロン酸に変換するHMG-CoA還元酵素を阻害すると、コレステロール合成の抑制だけでなく、増殖を促進するG結合タンパク質の活性化に必要なゲラニルグループやファルネシルグループの産生ができなくなる。Gタンパク質のイソプレニル化が阻害されるとがん細胞の増殖が抑制される。
細胞内におけるシグナル伝達においてGTP結合タンパク質(Gタンパク質)が重要な役割を担っています。
GTP結合タンパク質(Gタンパク質)は内在性のGTP加水分解(GTPase)活性をもつタンパク質の総称で、この内、低分子量Gタンパク質群(Ras, Rho,など)は分子量が2万~3万のタンパク質で、これまで100種類以上報告されています。RasやRhoはがん遺伝子として知られています。
イソプレニル化を受けた後に細胞膜に移行し、GTP結合型(on)/GDP結合型(off)として細胞内シグナル伝達に関与しています。
すなわち、Gタンパク質はGDP結合不活性型とGTP結合活性型の間をサイクル(GTPaseサイクル)することにより、細胞外からの情報を細胞内に伝達します。
Rhoは細胞骨格の制御を通して細胞の形態調節に関わっています。
低分子量Gタンパク質はそのほとんどでC末端から4番目のアミノ酸残基がシステインであり、ファルネシル基やゲラニルゲラニル基などの脂肪酸でファルネシル化されており、この脂質修飾により、細胞膜に直接結合できると考えられています。
HMG-CoA還元酵素を阻害してイソプレノイド生成が低下すると、低分子量Gタンパク質の活性が低下して、増殖活性が低下します。イソプレノイドはRasやRhoなどの低分子Gタンパク質が細胞膜の脂質に接着するときに必要が生体分子で、それができないと活性化できないので、がん細胞の増殖が抑制されることになります。

図:低分子量Gタンパク質の一種のRhoは、GDP結合型が不活性型で(①)、受容体などからの刺激を受けてGTP結合型となり、活性化され(②)、エフェクタータンパク質に作用して情報を伝達し、細胞の移動や細胞周期やアポトーシスや遺伝子発現を制御する(③)。HMG-CoAからメバロン酸経路で産生されるゲラニルゲラニルグループ(④)はRhoが細胞膜の脂質に接着するときに必要で、これができないとRhoは活性化できない(⑤)。HMG-CoAからメバロン酸を合成するHMG-CoA還元酵素の阻害剤であるスタチンはゲラニルゲラニルグループの産生を阻害することによってRhoの活性化を阻害する(⑥)。
【スタチンはイソプレノイド生成を低下してYAP/TAZの活性化を阻止する】
スタチンの抗がん作用については多くの基礎研究が行われており、スタチンによるメバロン酸経路の阻害は、がん細胞のシグナル伝達に影響し、がん細胞の増殖や転移の抑制、血管新生阻害、アポトーシス誘導などの抗がん作用が、培養細胞を使った実験や動物実験で報告されています。
メバロン酸経路でのイソプレノイド生成が低下するとYAP/TAZの活性化もできなくなります。
以下のような総説論文があります。
Metabolic control of YAP and TAZ by the mevalonate pathway.(メバロン酸経路によるYAPとTAZの代謝による制御)Nat Cell Biol. 2014 Apr;16(4):357-66.
【要旨】
Hippo経路のYAP/TAZは、組織の増殖および臓器の成長を促進する。しかしながら、それらの生物学的特性が細胞代謝とどのように関連するかは未解明のままである。
ここでは、YAP / TAZ活性がSREBP(Sterol regulatory element-binding protein) /メバロン酸経路によって制御されることを示す。
メバロン酸経路の律速酵素(HMG-CoAレダクターゼ)のスタチンによる阻害は、YAP / TAZの核局在および転写応答を阻害する。作用機序的には、メバロン酸経路によって産生されるゲラニルゲラニルピロリン酸はRho GTPアーゼの活性化に必要であり、YAP/TAZのリン酸化を阻害して核蓄積を促進することによってYAP / TAZを活性化する。
このメバロン酸とYAP / TAZの関係は、乳がん細胞の増殖および自己再生に必要である。
ショウジョウバエにおいては、メバロン酸生合成およびゲラニルゲラニル化の阻害は、YAP / TAZの相等体であるYorkieによって誘発された眼の過成長を阻止する。
腫瘍細胞においては、p53変異によって誘導されるSREBP転写活性によって産生されるメバロン酸のレベルの増加によってYAP / TAZ活性化は促進される。これらの知見は、代謝信号によるYAP / TAZ調節の存在を明らかにする。
転写アクチベーターのステロール調節エレメント結合タンパク質(The sterol regulatory element binding protein=SREBP)ファミリーは、コレステロールと脂肪酸の恒常性を調節する重要な制御因子です。
SREBPはコレステロール・脂肪酸代謝に関与する酵素遺伝子の発現を亢進します。
つまり、がん細胞では、SREBP転写活性によって産生されるメバロン酸のレベルの増加によってYAP / TAZ活性化は促進されます。したがって、メバロン酸経路を阻害するスタチンはYAP/TAZの活性化を阻止して、増殖抑制効果を発揮します。
しかし、スタチンによる抗腫瘍効果は薬剤の種類によって異なる可能性があります。つまり、スタチンには水溶性のものと脂溶性のものがあり、がん細胞に対する効果を期待するには脂溶性のものを使う必要があるようです。
水溶性スタチンは肝臓細胞膜に存在する有機アニオン輸送担体によって細胞内に取り込まれるので、肝細胞に選択的に取り込まれます。
脂溶性スタチンは細胞膜透過性が良いので、あらゆる臓器・組織の細胞内へ移行し得えます。
つまり、肝臓におけるコレステロール産生を抑制する目的では、他の細胞に影響が少ない点で水溶性スタチンの方が良いのですが、がん細胞に取り込まれて、メバロン酸経路を阻害して増殖抑制効果を期待するには脂溶性のものである必要があるようです。
スタチンの中で最も脂溶性の高いのがシンバスタチン(simvastatin)で、乳がんの患者さんがシンバスタチンを服用すると再発率が顕著に低下することが報告されています(548話参照)。
スタチンでメバロン酸経路を阻害すると体内でのCoQ10の産生が阻害されるので、CoQ10をサプリメントで補うのが良いと思います。
【COX-2阻害剤はYAP発現を抑制する】
シクロオキシゲナーゼ-2(COX-2)活性がYAPの活性を高めることが報告されています。つまり、COX-2阻害剤(celecoxibなど)はYAP活性の阻害に役立つ可能性があります。
以下のような報告があります。
COX-2 Forms Regulatory Loop with YAP to Promote Proliferation and Tumorigenesis of Hepatocellular Carcinoma Cells.(COX-2は、肝細胞がん細胞の増殖および腫瘍形成を促進するためのYAPとの調節ループを形成する。)Neoplasia. 2018 Apr;20(4):324-334.
【要旨】
シクロオキシゲナーゼ-2(COX-2)およびYAPは、肝細胞がんと高度に関連しており、肝細胞がんにおいてCOX-2とYAPの高発現が高い頻度で認められる。 しかしながら、COX-2とYAPの間に相互作用があるかどうか、そしてそれらが互いにどのように相互作用するかは、重要ではあるが明確ではない。
この論文では、我々は、HCC細胞株におけるCOX-2の過剰発現が、YAPの mRNAとタンパク質、およびその標的遺伝子の発現レベルの増加をもたらすことを示した。
COX-2はHCC細胞株の増殖を促進し、YAP遺伝子の欠損はこの効果を阻止した。 さらに、我々の結果は、EP2(prostaglandin E-receptor 2)およびWnt /βカテニンがCOX-2によるYAPの転写誘導を媒介することを示した。 一方、YAPは、COX-2プロモーターにおけるTEAD結合部位を介した転写レベルでCOX-2発現を増加させた。
まとめると、これらの所見は、COX-2がYAPを活性化するだけでなく、Hippo-YAP経路の標的でもあり、したがってCOX-2-PGE2-EP2-Gαs-βカテニン-YAP-COX -2というフィードバック回路を形成している。
さらに、YAとCOX-2の同時阻害は、いずれか単独の阻害よりも相乗的かつより効率的に細胞増殖と腫瘍形成を低下させた。これはYAPおよびCOX-2のフィードバック回路を遮断することが肝臓がんの有効な治療法になることを示唆している。
つまり、COX-2はYAPシグナル系を活性化することによって肝臓がんの発生に関与している可能性を示唆しています。
さらに、COX-2はWnt/β-cateninを介してYAPのmRNAとタンパク質レベルを高めることを示しています。COX-2によるYAP発現はβカテニン欠損細胞では認められませんでした。
さらに、COX-2の遺伝子にはTEAD結合部位がありYAPによって発現が亢進することが示されています。
つまり、COX-2とWnt/β-cateninとYAPは相互に作用してポジティブなフィードバック回路を形成して、増殖促進作用を強めていることを明らかにしています。
この回路が肝臓がんの発生と進展を促進している可能性を示唆しています。βカテニンの活性を阻害する方法はYAP発現抑制に寄与する可能性があります。
つまり、COX-2阻害剤(セレコックス)と、Wint/βカテニン阻害剤とYAP阻害剤(シンバスタチン)を組み合せると肝臓がんの治療効果を高めることができるかもしれません。Wint/βカテニン阻害剤としてはビタミンD3とスリンダクが使えます(462話参照)。
| « 593) 若い女性... | 595) Hippo経... » |