がんの予防や治療における漢方治療の存在意義を考察しています。がん治療に役立つ情報も紹介しています。
「漢方がん治療」を考える
カレンダー
| 2025年1月 | ||||||||
| 日 | 月 | 火 | 水 | 木 | 金 | 土 | ||
| 1 | 2 | 3 | 4 | |||||
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | ||
| 12 | 13 | 14 | 15 | 16 | 17 | 18 | ||
| 19 | 20 | 21 | 22 | 23 | 24 | 25 | ||
| 26 | 27 | 28 | 29 | 30 | 31 | |||
|
||||||||
goo ブログ
過去の記事
カテゴリ
最新の投稿
最新のコメント
最新のトラックバック
ブックマーク
プロフィール
検索
gooおすすめリンク




| URLをメールで送信する | |
| (for PC & MOBILE) | |
869)抗がん剤治療はなぜ失敗するのか(その2): アポトーシス細胞ががん幹細胞の再増殖を促進するフェニックス・ライジング(不死鳥復活)
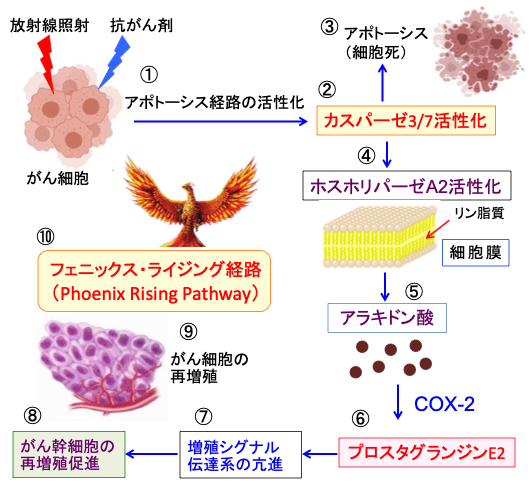
図:がん細胞が放射線照射や抗がん剤で強いダメージを受けるとアポトーシス経路が活性化され(①)、最終的にアポトーシスを実行するカスパーゼ3と7が活性化されて(②)、細胞死(アポトーシス)が誘導される(③)。活性化したカスパーゼ3と7は細胞のホスホリパーゼA2を切断して活性化し(④)、細胞膜のリン脂質からアラキドン酸を産生し(⑤)、がん細胞に過剰に発現しているシクロオキシゲナーゼ-2(COX-2)によってプロスタグランジンE2が産生され、細胞外に分泌される(⑥)。プロスタグランジンE2はがん細胞の増殖シグナル伝達系を刺激して、細胞増殖を亢進する作用がある(⑦)。そのため、プロスタグランジンE2によって、生き残ったがん幹細胞の再増殖が促進され(⑧)、がん組織は増大する(⑨)。アポトーシスによる細胞死を起こしたがん細胞によって生き残ったがん幹細胞の再増殖が促進される経路はフェニックス・ライジング(不死鳥復活)経路と呼ばれている(⑩)。
869)抗がん剤治療はなぜ失敗するのか(その2): アポトーシス細胞ががん幹細胞の再増殖を促進するフェニックス・ライジング(不死鳥復活)
【アポトーシスは炎症反応を起こさない細胞死】
細胞死のパターンはいくつかありますが、代表的なのが壊死(ネクローシス)とアポトーシスです。
脳梗塞や心筋梗塞の様な虚血や、火傷や毒物による細胞傷害では、壊死(ネクローシス)という細胞死を起こして、細胞が崩壊して炎症反応が引き起こされます。
外部からの急激なダメージ(虚血、低酸素、外傷など)によって引き起こされる非計画的な細胞死と言えます。細胞内容物の漏出が炎症反応を引き起こします。
炎症反応を引き起こすことによって、生体に異常事態を知らせ、防御や修復を促進することができます。
アポトーシスは、細胞のプログラムに従った、自律的な「自殺」のような形で起こる計画的な細胞死です。形態学上は核の凝集と細胞収縮、それにDNAの断片化などによって特徴づけられます。
細胞は収縮し、細胞の断片(アポトーシス小体)が形成され、最終的にマクロファージなどの貪食細胞によって取り込まれて処理されるので、基本的には炎症反応を引き起こしません。

図:壊死(ネクローシス)は外部からの急激なダメージ(外傷、虚血、低酸素など)によって引き起こされる細胞死で、細胞が膨張し、細胞膜が破れ、細胞内容物が周囲の組織に漏れ出す。細胞内容物の漏出が炎症反応を引き起こす。一方アポトーシスは、細胞のプログラムに従った、自律的な「自殺」のような形で起こる計画的な細胞死で、細胞は収縮し、細胞の断片(アポトーシス小体)が形成され、最終的にマクロファージなどの貪食細胞によって取り込まれる。アポトーシスは炎症を引き起こさない。
生物はその発生や成長過程で、最初に余分に作成して、不要なものを削っていくという方法を採用しています。
例えば、発生の過程で、指ができるとき、まず細胞の塊ができ、その細胞の塊の一部がアポトーシス(プログラム細胞死)で削られて指ができます。このようにして手が形成されます。(下図)

図:生物は発生や成長の過程でまず過剰に作って、不要なものを削って完成させるという方法を採用する。例えば、手の発生過程では、手の原型となる細胞の塊の一部がアポトーシス(プログラム細胞死)によって削られて、指を作って手が完成する
このように、アポトーシスは、個体の発生段階で過剰にできた細胞を選択的に除去し、器官形成を完成するのに必要なプロセスです。
また、老化した細胞や修復不能なダメージを受けた細胞を除去する際にも、アポトーシスのメカニズムで細胞死が起こります。
細胞は軽度のダメージ(傷害)の場合は、ダメージを修復して生き残ります。ダメージが強い場合は、不完全な修復では異常細胞(がん細胞や老化細胞)になるので、自滅する道を自ら選択します。この時に実行される細胞死もアポトーシスです。遺伝子に大きなダメージを受けたとき、がん細胞の発生を防ぐメカニズムになります。
アポトーシスの過程では、細胞の体積が縮小し、細胞核が縮んで、染色体が断片化します。さらに、細胞の断片、いわゆるアポトーシス小体が形成され、周囲の細胞やマクロファージによって迅速に取り込まれ消化されます。(下図)
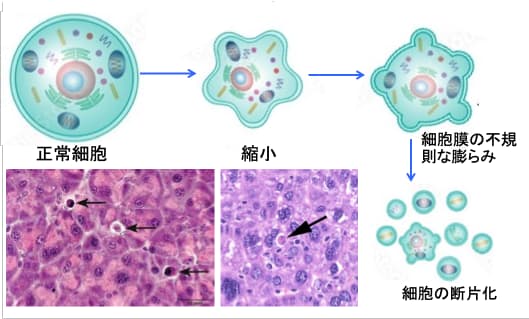
図:アポトーシスでは、細胞は縮小し、細胞膜の不規則な膨らみができ、細胞が断片化される。断片化したアポトーシス小体は、周囲の細胞やマクロファージなどによって処理される。病理組織の矢印がアポトーシス細胞。
細胞が崩壊して細胞内成分が漏出すると炎症反応が惹起されます。生理的な細胞死に対して体が炎症反応を起こさないようにするために細胞が収縮し、断片化されて周囲の細胞やマクロファージなどによって迅速に処理されるのがアポトーシスの特徴です。炎症を誘発せず、周囲の組織や細胞にできるだけ迷惑をかけないで細胞交代型の細胞死です。
【アポトーシスではカスパーゼというタンパク質分解酵素が活性化されて細胞死を実行する】
細胞のミトコンドリアは細胞が必要とするエネルギー(ATP)の大部分を産生しています。さらに、カルシウム恒常性の維持、ヘム合成、栄養素の代謝、ステロイドホルモンの合成、尿素合成によるアンモニアの除去、様々な物質代謝、アポトーシスやオートファジーのシグナル伝達系の制御など様々な機能を持っています。
アポトーシス実行過程では、ミトコンドリアに存在するたんぱく質が重要な役割を果たしています。
たとえば、電子伝達系で働くシトクロムCというたんぱく質は、ミトコンドリアから放出されると、たんぱく分解酵素群が次々に活性化されて、細胞死へと導きます。
アポトーシスは多数のタンパク質によって制御されていますが、中心になるのがBcl-2やBaxと呼ばれるタンパク質群(Bcl-2ファミリー蛋白質)です。
Bcl-2 ファミリー蛋白質には、アポトーシスを抑制するBcl-2-サブファミリー(Bcl-2, Bcl-Xlなど)と、アポトーシスを促進する Baxサブファミリー(Bax、Bakなど) および BH3-onlyサブファミリー (Bid、Bim、Badなど)があります
Bcl-2およびそのファミリー蛋白質の主な作用部位はミトコンドリア膜で、このミトコンドリア膜の透過性を制御することにより、細胞の生死を決定しています。
ミトコンドリアの膜透過性が亢進すると、膜間スペースに存在するアポトーシス誘導蛋白質(シトクロムcなど)が細胞質に漏出します。漏出したシトクロムcはカスパーゼと呼ばれる蛋白分解酵素群を段階的に活性化してアポトーシスが実行されます。
Baxはアポトーシス実行時のミトコンドリアの膜透過性亢進に必須の分子として働き、Bcl-2はBaxと直接結合してBaxの働きを阻止します。
Bcl-2のようなアポトーシス促進蛋白質の活性が、Baxのような抑制蛋白質の活性を凌駕した場合に、ミトコンドリア膜透過性亢進が誘導され、アポトーシスが実行されます。
Bcl-2サブファミリー蛋白質(Bcl-2やBcl-xL)を過剰発現したがん細胞は多くのアポトーシス刺激(抗がん剤治療など)に対して強い抵抗性を示します。したがって、Bcl-2サブファミリー蛋白質の活性や発現量を抑制したり、Baxサブファミリー蛋白質の活性や発現量を高める効果は、がん細胞の抗がん剤感受性を高め、死滅しやすくなります。

図:Bcl-2はBaxと結合してBaxの活性を抑制している。抗がん剤や放射線照射などによって細胞を傷害されるとBH3ドメインをもつ蛋白質がBcl-2に結合してBcl-2を不活性化する。その結果フリーになったBaxはミトコンドリア外膜にて二量体(ダイマー)になって内腔を形成してミトコンドリアからシトクロム C を含む様々なタンパク質が細胞質内に流出し、蛋白分解酵素のカスパーゼ(Caspases)が活性化されアポトーシスが実行される。
アポトーシス(Apoptosis)はミトコンドリアのチトクロムCやタンパク分解酵素のカスパーゼが関与する細胞死です。カスパーゼは、基質であるタンパク質をアスパラギン酸残基の後で切断するシステインプロテアーゼの総称です。アポトーシスの実行や炎症性サイトカインの活性化に関与するタンパク分解酵素です。
カスパーゼは初め不活性型の前駆体のプロ・カスパーゼ(pro-Caspase)として発現しますが、Fas などの細胞死受容体や各種の傷害刺激などに反応して速やかに切断され、活性型となります。
アポトーシスの過程においては、誘導型カスパーゼ(Initiator caspase)と実行型カスパーゼ(Executioner caspase / Effector caspase)に大別されます。
誘導型カスパーゼ(Caspase-2、8、9、10)は実行型カスパーゼ(Caspase-3、6、7)を切断・活性化することによりアポトーシスのシグナルをスタートさせます。実行型カスパーゼは、多くの細胞内タンパク質を分解し、アポトーシスの実行へと導きます(下図)。

図:細胞死受容体(Fas と TNFR)のリガンド(FasLとTNF)による活性化(外因性経路)は、カスパーゼ8 および カスパーゼ10を活性化する(①)。ダメージを受けたミトコンドリアから放出されるシトクロムC はカスパーゼ-9 を活性化する(内因性経路②)。これらのカスパーゼはイニシエーターカスパーゼ(Initiator caspase)と呼ばれ、下流のエクセキューショナーカスパーゼ(Executioner caspase)のpro-caspase3とpro-caspase 7を切断して活性化する(③)。活性化したカスパーゼ3とカスパーゼ7は細胞内タンパク質を消化してアポトーシスを実行する。
【アポトーシス細胞が増殖シグナルを残す】
アポトーシスは、細胞が収縮・断片化し、マクロファージなどの貪食細胞や周辺の細胞によって取り込まれます。炎症を引き起こさないで、周囲細胞に気づかれずに死滅するような印象があります。
しかし実際は、アポトーシス細胞は残った細胞の増殖を刺激するようなシグナルを残すことが知られています。
例えば、アポトーシスが進行すると、細胞からさまざまなサイトカインや成長因子が放出されることがあります。これらの分子は、隣接する細胞にシグナルを送り、これにより細胞の増殖、移動、または分化が刺激されることがあります。
さらに、アポトーシスする細胞は、エクソソームと呼ばれる小さな微小泡を放出することが知られています。これらのエクソソームは、RNA、蛋白質、脂質などの様々な分子を運び、隣接する細胞との間で情報の交換を助けることができます。このコミュニケーションは、近隣の細胞の行動を変え、増殖を刺激する可能性があります。
これらの機構を通じて、アポトーシスは組織の再生や機能の維持をサポートする役割を果たします。
創傷治癒と組織再生は、すべての多細胞生物にとって不可欠なプロセスです。常在する組織幹細胞は、創傷治癒と組織再生において重要な役割を果たします。損傷した組織は幹細胞を動員して損傷部位に補充し、そこで増殖、分化し、最終的には損傷した組織を置き換えます。
組織損傷に応じた組織再生のトリガーとしてアポトーシス実行過程でのカスパーゼ活性化が細胞増殖と幹細胞動員の重要な要件であることが示されています。
一般に、アポトーシスは多細胞生物が損傷した細胞や不要な細胞を処分するための手段であると考えられているため、アポトーシス細胞が組織の修復に積極的に関与するという考えは素直に納得できない感じもあります。
しかし、死にかけている細胞がその存在を周囲の組織にシグナル伝達し、そうすることで組織の修復と再生を引き起こし、それを補う可能性があるという証拠が増えています。哺乳類モデルにおける最初の証拠は、2010年に発表され、組織の損傷が組織の修復を開始する「フェニックス・ライジング経路(Phoenix Rising Pathway)」というメカニズムが提唱されました。
【創傷治癒を促進するフェニックス・ライジング(Phoenix Rising)経路】
フェニックス(Phoenix)というのは、死んでも蘇ることで永遠の時を生きるといわれる伝説上の鳥です。500~600年程生きた後、自ら火中に入って焼かれて死にますが、その灰の中から幼鳥の姿となって再び蘇って永遠の時を生きるとされています。「不死鳥」や「火の鳥」とも呼ばれます。

フェニックス・ライジング(Phoenix Rising)というのは「不死鳥復活」という意味です。
創傷治癒に関与するシグナル伝達系にも「Phoenix Rising Pathway」というシグナル伝達系が提唱されています。以下のような論文があります。米国のコロラド大学医学部の放射線腫瘍学部門などの研究グループからの論文です。
Apoptotic cells activate the "phoenix rising" pathway to promote wound healing and tissue regeneration.(アポトーシス細胞がフェニックス・ライジング経路を活性化し、創傷治癒と組織再生を促進する)Sci Signal. 2010 Feb 23;3(110):ra13. doi: 10.1126/scisignal.2000634.
【要旨】
損傷を受けた組織を再生する能力は、多細胞生物に共通する特徴である。我々は、マウスの創傷治癒と組織再生を促進する際にアポトーシス細胞が果たす役割について報告する。
アポトーシス細胞は、前駆細胞(幹細胞から最終分化細胞へと分化する途中の細胞)または幹細胞の増殖を促進する増殖シグナルを放出した。この過程における重要なタンパク質は、細胞死(アポトーシス)の実行段階に活性化されるプロテアーゼ(タンパク分解酵素)のカスパーゼ3とカスパーゼ7であった。
この2つのカスパーゼのいずれか一方を欠失したマウスでは、皮膚の創傷治癒と肝臓再生が不十分であった。
幹細胞または前駆細胞の増殖と組織再生の促進因子であるプロスタグランジンE2は、この2つのカスパーゼの下流で作用した。
我々は、アポトーシスの実行者であるこれらのカスパーゼが多細胞生物における創傷治癒と組織再生を促進する経路を、フェニックス・ライジング(phoenix rising)経路と呼ぶことを提唱する。
組織や臓器が損傷を受けると、残った細胞が増殖して、損傷を修復するメカニズムが作動します。
創傷治癒や組織再生は、多くの細胞が協調的に働く極めて複雑なプロセスです。
一般的には、炎症細胞が組織傷害部位の最初の応答者であると考えられています。マクロファージや好中球のような炎症細胞が、創傷治癒や組織再生プロセスの開始と進行の制御において決定的に重要であるという考えが長くありました。
しかし、マクロファージおよび好中球を欠くノックアウトマウスでも創傷治癒は正常に行われるので、いずれの細胞タイプも皮膚切除創傷治癒には必要でないことが示されています。
損傷した組織の中および周囲の幹細胞は、創傷治癒および組織再生において重要な役割を果たしています。
一般に、損傷組織から放出された因子が、これらの幹細胞および前駆細胞を動員して傷害部位に補充し、それらが増殖し、分化し、最終的に損傷組織を再生すると考えられています。
組織損傷から生じる炎症過程の間に産生される様々な炎症性サイトカインや増殖因子が、創傷治癒や組織再生のシグナル伝達に関与しています。
この論文では、傷ついた組織の死細胞(アポトーシス細胞)が、組織再生および創傷治癒のプロセスを開始する幹細胞または前駆細胞(幹細胞から最終分化細胞へと分化する途中の細胞)の増殖を刺激するシグナルを送信することを示し、フェニックス・ライジング(phoenix rising)経路と呼ぶことを提唱しています。
【カスパーゼ3および7の遺伝子欠損マウスでは創傷治癒過程が阻害される】
カスパーゼは、基質であるタンパク質をアスパラギン酸残基の後で切断するシステインプロテアーゼの総称です。アポトーシスの実行や炎症性サイトカインの活性化に関与するタンパク分解酵素です。
アポトーシスの過程においては、誘導型カスパーゼ(Initiator caspase)と実行型カスパーゼ(Executioner caspase/Effector caspase)に大別されます。
誘導型カスパーゼ(Caspase-2、8、9、10)は実行型カスパーゼ(Caspase-3、6、7)を切断・活性化することによりアポトーシスのシグナルをスタートさせます。実行型カスパーゼは、多くの細胞内タンパク質を分解し、アポトーシスの実行へと導きます。
カスパーゼ3および7の遺伝子欠損マウスでは創傷治癒過程が阻害されることが報告されています。つまり、創傷を起こしても、幹細胞と前駆細胞(幹細胞から最終分化細胞へと分化する途中の細胞)の増殖を刺激しないのです。
そこで、先ほどの論文では、アポトーシスを実行する際に最終的に作用するカスパーゼ3と7が、組織幹細胞の増殖を促進するメカニズムが検討されています。
アポトーシスを起こした細胞の中で、カスパーゼ3とカスパーゼ7がカルシウム非依存性ホスホリパーゼA2(calcium-independent phospholipase A2 :iPLA2)を活性化して脂肪膜のリン脂質からアラキドン酸の合成を亢進し、さらにシクロオキシゲナーゼの作用によってプロスタグランジンE2が産生されます。
プロスタグランジンE2は組織幹細胞の増殖を刺激し、組織の再生や創傷治癒過程を促進する作用があります。
カスパーゼが正常な細胞に放射線照射を行うとアラキドン酸の産生が亢進しますが、カスパーゼ3の遺伝子欠損細胞に放射線照射を行ってもアラキドン酸は増えませんでした。
カルシウム非依存性ホスホリパーゼA2(iPLA2)は活性化したカスパーゼ3によって切断されることによって活性化することが示されています。
プロスタグランジン(Prostaglandins; PGs)はアラキドン酸からシクロオキシゲナーゼ(Cyclooxygenase; COX)の働きにより合成される生理活性物質で、炎症の代表的なメディエーターです。PGsはいわゆる「アラキドン酸カスケード」によって産生されます。
すなわち、細胞外から種々の刺激に反応して生体膜のリン脂質がホスホリパーゼA2 (PLA2)により、まず不飽和脂肪酸のアラキドン酸に変換されます。この遊離したアラキドン酸を基質として、脂肪酸酸化酵素であるシクロオキシゲナーゼ(COX)の作用により、PGG2, PGH2へと変換され、さらに各種細胞に存在する特異的な合成酵素により生理的に重要な4種類のPGs (PGD2, PGE2, PGF2a, PGI2)とトロンボキサン(thromboxane; TX)A2が合成されます。
COXにはCOX-1とCOX-2の2種類のアイソザイムが知られています。この2つのCOXは約60%のアミノ酸配列の相同性をもっていますが、それぞれ生体内での役割が異なることが明らかになっています。
COX-1は胃や腸などの消化管、腎臓、卵巣、精嚢、血小板などに存在し、胃液分泌、利尿、血小板凝集などの生理的な役割を担います。一方、COX-2はサイトカインや発がんプロモーター、ホルモンなどの刺激により、マクロファージ、線維芽細胞、血管内皮細胞、癌細胞などで誘導され、炎症反応、血管新生、アポトーシス、発癌、排卵、分娩、骨吸収などに関与しています(下図)

図:ホスホリパーゼA2(PLA2)の働きで、細胞膜のリン脂質からアラキドン酸が生成される。シクロオキシゲナーゼ(COX)はアラキドン酸からプロスタグランジンを合成するときに最初に働く酵素で、COX-1とCOX-2の2種類がある。COX-1から合成されるプロスタグランジンは生体の生理機能に必要なものであるが、炎症性の刺激でCOX-2から合成される大量のプロスタグランジンは細胞の増殖を促進する。
アポトーシスを起こしている細胞内で、実行型カスパーゼのカスパーゼ3と7がホスホリパーゼA2を切断して活性化し、アラキドン酸を産生し、さらにシクロオキシゲナーゼ1および2によってプロスタグランジンE2が産生され、このプロスタグランジンE2が組織幹細胞の増殖を促進し、創傷治癒過程を開始し促進するというメカニズムを明らかにして、この経路をフェニックス・ライジング(不死鳥復活)経路と呼ぶことを提唱しています。この論文のまとめの図を以下の示しています。

図:損傷を受けた組織(①)では、アポトーシスを起こしている細胞内では、カスパーゼ9が関与する内因性経路やカスパーゼ8が関与する外因性経路(②)によって、カスパーゼ3とカスパーゼ7が活性化され(③)、アポトーシスが実行される。活性化したカスパーゼ3と7は不活性型のホスホリパーゼA2(PLA2)を切断して活性型にする(④)。活性化したPLA2は細胞膜のリン脂質からアラキドン酸を産生する(⑤)。アラキドン酸はシクロオキシゲナーゼ-1と2(COX-1とCOX-2)によってプロスタグランジンH2(PGH2)に変換され(⑥)、PGH2はプロスタグランジンE2(PGE2)合成酵素によってPGE2が産生される(⑦)。産生されたPGE2は組織幹細胞の増殖を促進して、正常組織の再生を進める(⑧)。
【放射線治療や抗がん剤治療でアポトーシスを起こしたがん細胞はがん幹細胞の再増殖を刺激する】
正常組織では、フェニックス・ライジング経路は損傷組織の修復や再生を促進します。
がん組織では、この経路は放射線治療や抗がん剤治療後の生き残ったがん細胞の再増殖を促進するように作用します。
正常組織もがん組織も、アポトーシス細胞は残った幹細胞の増殖を促進するシグナルを残して死ぬというのがフェニックス・ライジング経路なのですが、この経路の存在はがん治療が失敗する原因にもなります。
放射線治療や抗がん剤治療や放射線治療によってDNAがダメージを受けると、がん細胞は細胞分裂を中止し、ダメージを受けたDNAの修復を開始します。
しかし、ダメージが修復不可能なレベルに大きいと、アポトーシスが実行されます。
このアポトーシスの実行の最終段階で働くのがカスパーゼ3と7です。
このカスパーゼはホスホリパーゼA2を切断して活性化し、アラキドン酸を合成して、さらにPGE2の産生を増やします。がん組織ではがん細胞や炎症細胞がCOX-2を過剰に発現しているので、PGE2の産生量は正常組織より高度になります。
PGE2は生き残ったがん細胞の増殖を促進する増殖因子になります。特に、がん幹細胞の増殖を促進します。
つまり、ダメージを受けたがん細胞はアポトーシスを起こすときに、生き残ったがん幹細胞の増殖活性を高めるようなシグナルを残して死ぬのです。
培養しているがん細胞に、放射線照射してアポトーシスを起こしているがん細胞を加える実験があります。カスパーゼ3/7を持っているがん細胞を放射線でアポトーシスを誘導すると、がん細胞の増殖が促進されます。しかし、カスパーゼ3/7が欠損しているがん細胞を放射線で死滅しても、他のがん細胞の増殖を促進しません。
つまり、アポトーシスを起こしている細胞で活性化されるカスパーゼ3/7の存在が、がん細胞の増殖を促進するということです。
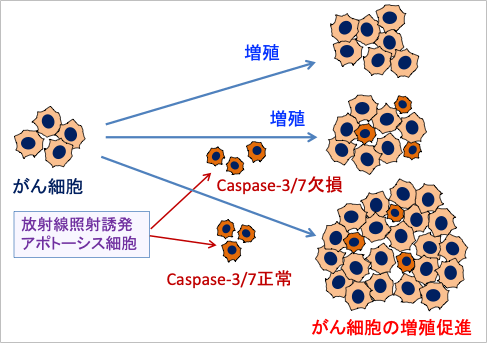
図:培養しているがん細胞に放射線照射で誘発したアポトーシス細胞を混ぜて培養する実験で、Caspase-3(カスパーゼ3)とCaspase-7が正常な細胞のアポトーシスの場合は、がん細胞の増殖を促進する。しかし、Caspase-3/7遺伝子を欠損しているアポトーシス細胞では、がん細胞の増殖を促進しない。これは放射線でアポトーシスを起こしている細胞のCaspase-3/7活性ががん細胞の増殖を促進することを意味している。
そこで、放射線治療や抗がん剤治療を行うときにCOX-2阻害剤のセレコキシブ(celecoxib)を併用するとプロスタグランジンE2の産生を抑制して、がん幹細胞の再増殖を阻止できる可能性が指摘されています。
以下のような報告があります。フェニックス・ライジング経路を最初に提唱した研究者からの総説論文です。
Cyclooxygenase-2: A Role in Cancer Stem Cell Survival and Repopulation of Cancer Cells during Therapy(シクロオキシゲナーゼ-2:がん治療中のがん幹細胞の生存とがん細胞の再増殖における役割)Stem Cells Int. 2016; 2016: 2048731.
【要旨】
シクロオキシゲナーゼ-2(COX-2)は、プロスタグランジンE2を含むプロスタグランジン類を合成する誘導型の酵素である。プロスタグランジンE2は炎症や血管新生の主要な化学伝達物質として働く。
COX-2はがん細胞において過剰に発現しており、がん細胞の進行性の増殖と、抗がん剤治療や放射線治療に対するがん細胞の抵抗性の要因になっている。
抗がん剤治療も放射線治療も多数の回数に分けて投与される。これは治療の間にダメージを受けた正常細胞を回復させるためである。
しかし、がん治療を中断している間に生き残ったがん細胞も増殖し、腫瘍の再増殖を引き起こし、治療の有効性を低下させている。このようながん細胞の再増殖が治療の失敗の主な原因になっている。
通常の抗がん剤治療や放射線治療では、抵抗性の高いがん細胞が選択的に生き残るので、がん細胞の再増殖が起こる。
COX-2の過剰発現とプロスタグランジンE2の産生と放出が、腫瘍の再増殖と患者の予後不良の原因となっていることを示す十分なエビデンスが存在する。
この総説では、がん幹細胞の生物学におけるCOX-2の役割と、がん細胞の再増殖のメディエーターとしての役割を考察し、がん治療に対する抵抗性を克服するターゲットになることを示す。
抗がん剤でがん組織が縮小しても、多くは一時的で、しばらくして再増殖し、転移します。
組織のダメージが組織修復を開始するシグナルになるとき、フェニックス・ライジング経路というシグナル伝達系が働き、その際にプルスタグランジンE2(PGE2)が重要な役割を担っています。
放射線照射やDNA傷害性の抗がん剤ががん細胞にアポトーシスを誘導するとき、アポトーシスを起こしている細胞からPGE2が放出されます。このPGE2は強力な増殖因子となって、生き残ったがん幹細胞の増殖を刺激します。その結果、がん細胞の再増殖が起こります。
つまり、PGE2の産生を増やすCOX-2ががんの再増殖や再発を促進するのです。
図:放射線照射や抗がん剤で損傷を受けたがん細胞(①)は、カスパーゼ9が関与する内因性経路やカスパーゼ8が関与する外因性経路(②)によって、カスパーゼ3とカスパーゼ7が活性化され(③)、アポトーシスが実行される。活性化したカスパーゼ3と7は不活性型のホスホリパーゼA2(PLA2)を切断して活性型にする(④)。活性化したPLA2は細胞膜のリン脂質からアラキドン酸を産生する(⑤)。アラキドン酸はシクロオキシゲナーゼ-1と2(COX-1とCOX-2)によってプロスタグランジンH2(PGH2)に変換され(⑥)、PGH2はプロスタグランジンE2(PGE2)合成酵素によってPGE2が産生される(⑦)。産生されたPGE2はがん幹細胞の増殖を促進して、がん細胞の再増殖を促進する(⑧)。
したがって、COX-2の選択的阻害剤のセレコキシブ(celecoxib)は放射線治療や抗がん剤治療によって引き起こされるがん幹細胞の再増殖を抑制する効果が期待できます。
メトホルミンもCOX2/PGE2/STAT3経路を阻害する機序でフェニックス・ライジング経路を阻害できる可能性があります。以下のような報告があります。
Metformin represses bladder cancer progression by inhibiting stem cell repopulation via COX2/PGE2/STAT3 axis(メトホルミンはCOX2/PGE2/STAT3経路を介するがん幹細胞の再増殖を阻害することによって膀胱がんの進行を抑制する)Oncotarget. 2016 May 10; 7(19): 28235–28246.
【要旨】
がん幹細胞は、膀胱がんを含む様々ながんの発生、分化、再発、転移および薬剤耐性の獲得に必須の役割を果たす腫瘍細胞の亜集団である。
異なるがん種においてメトホルミンががん幹細胞の再増殖を抑制することが、多くの研究によって示されている。しかし、メトホルミンの膀胱がん幹細胞に対する効果はほとんど分かっていない。
我々は、N-メチル-N-ニトロソ尿素(MNU)誘発ラット膀胱がんモデルを使用して、メトホルミンが軽度から中等度/重度の異形成病変および上皮内がんから浸潤性のがんに移行するのを抑制することを明らかにした。
さらに、メトホルミンが、膀胱がん細胞をG1 / S期に停止させ、アポトーシスを誘導することを示した。
また、メトホルミンは、動物実験モデルおよび培養細胞モデルの両方においてがん幹細胞のマーカーであるサイトケラチン14(CK14 +)およびoctamer-binding transcription factor 3/4 (OCT3/4+)陽性の細胞を減少させることによって、膀胱がん幹細胞の再増殖を抑制することが示された。
さらに重要なことに、我々は、メトホルミンがCOX2活性を阻害し、PGE2産生ならびにSTAT3の活性化を阻害することによって、これらの抗がん効果を発揮することを見出した。
結論として、我々は、メトホルミンがCOX2 / PGE2 / STAT3軸を介したがん幹細胞再増殖を阻害することにより、膀胱がんの進行を阻害することを動物モデルおよび培養細胞モデルの両方において初めて系統的に立証した。
以上の報告から、抗がん剤治療や放射線治療後に発生するがん細胞の再増殖を抑える方法としてCOX-2阻害剤のセレコキシブのほかにPGE2の産生を抑制するメトホルミン、ドコサヘキサエン酸の併用が有効と言えます。(下図)

図:がん細胞が放射線照射や抗がん剤で強いダメージを受けるとアポトーシス経路が活性化され(①)、最終的にアポトーシスを実行するカスパーゼ3と7が活性化されて(②)、細胞死(アポトーシス)が誘導される(③)。活性化したカスパーゼ3と7は細胞のホスホリパーゼA2を切断して活性化し(④)、細胞膜のリン脂質からアラキドン酸を産生し(⑤)、がん細胞に過剰に発現しているシクロオキシゲナーゼ-2(COX-2)によってプロスタグランジンE2が産生され、細胞外に分泌される(⑥)。プロスタグランジンE2はがん細胞の増殖シグナル伝達系を刺激して亢進する作用がある(⑦)。そのため、プロスタグランジンE2によって、生き残ったがん幹細胞の再増殖が促進され、がん組織は増大する(⑧)。アポトーシスによる細胞死を起こしたがん細胞によって生き残ったがん幹細胞の再増殖が促進される経路はフェニックス・ライジング(不死鳥復活)経路と呼ばれている。COX-2の阻害剤であるセレコキシブ(celecoxib)、メトホルミン、DHA(ドコサヘキサエン酸)はフェニックス・ライジング経路を阻害して、放射線治療や抗がん剤治療後の再増殖を阻止する(⑩)。
セレコキシブもメトホルミンも極めて安価で安全性の高い薬です。ドコサヘキサエン酸は食品です。抗がん剤や放射線治療と併用しても、副作用を増悪する可能性は低い薬です。抗がん剤治療と放射線治療後のがん細胞の再増殖を抑える効果を示すエビデンスは高いと言えます。がん治療中の補完医療として試してみる価値はかなり高いと思います。
新刊紹介

抗がん剤治療が失敗する理由を解説し、その原因に対処する補完医療についても解説しています。さらに、抗がん剤治療の止め時を適切に判断し、終末期における「生活の質」と「死の質」の両方を良くすることの大切さを解説しました。死を早める可能性もある「無駄な抗がん剤治療」を避けるためには、医師の言いなりにならずに、患者自身が正しい知識を得て、もっと考える必要があると思います。
| « 868)抗がん剤... | 870)抗がん剤... » |