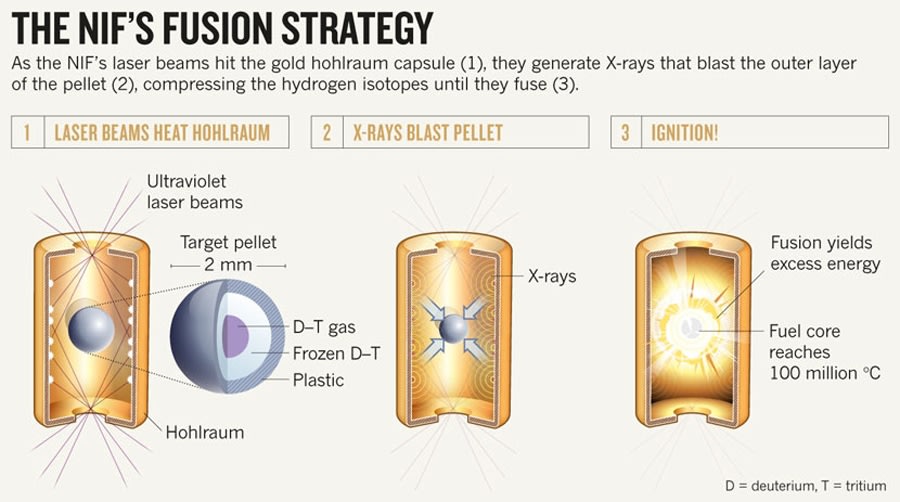
【技術革新情報とその処理】
このブログをはじめて5年になるが、情報処理術も少しは上達したこともあり、検索速度の向上
により膨大な量になりそれに伴う眼精脳疲労の負荷も逓増するが、問題は情報量ではなく、その
質にある。といっても、その情報の価値を推し図り、仕分けストレージするのも自分自身である。
特に、科学技術向上の情報収集は半端じゃなくめざましい昨今だと実感する(『爆発する技術革
新』)。今夜はその一例を考えてみる。その上で、発展著しい再生医療をはじめとした生物工学
(『成熟に向かう新弥生時代(2)』)の研究開発事例を俯瞰し、その大本の”生物の起源”について
考えてみる。
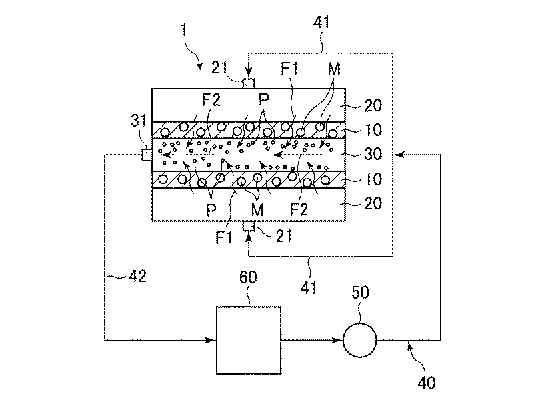
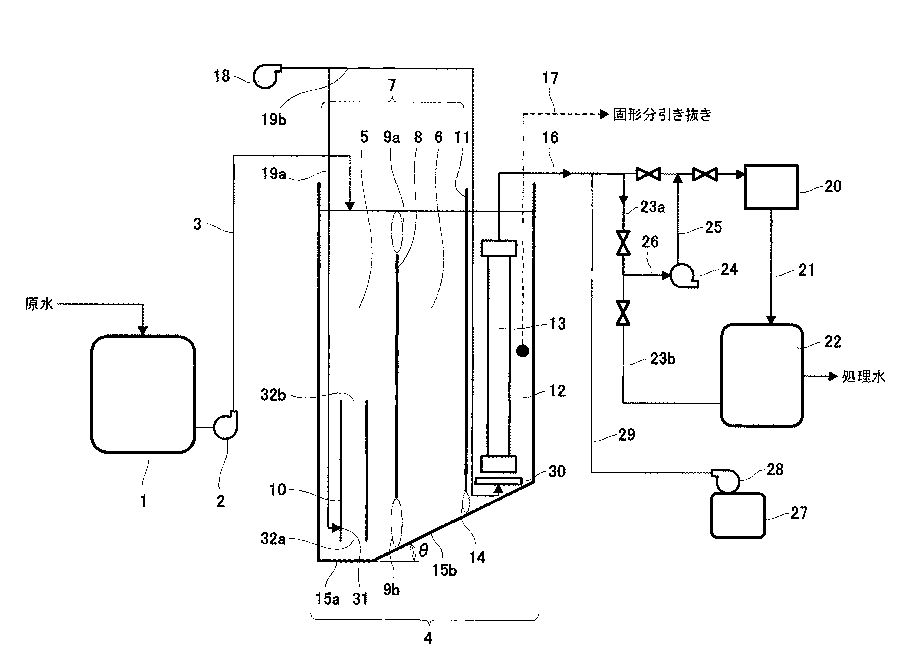
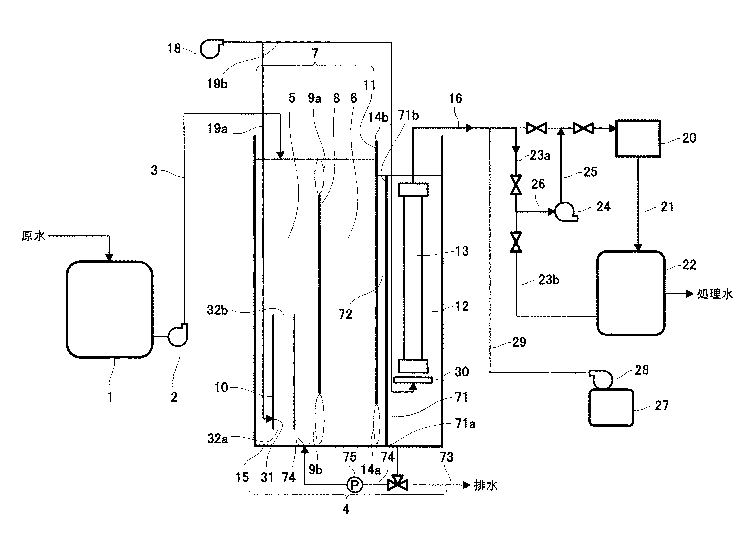
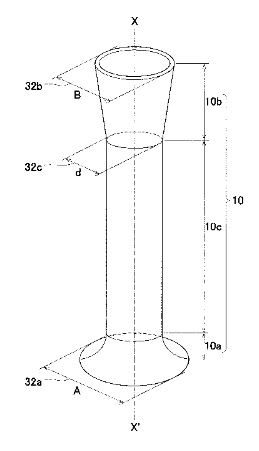
●微粒子を密度差により簡便に分別する

従来、細胞や化学合成された粒子等の、粒子毎に異なる性質の粒子群を分別する方法として、質
量差により分別する方法、大きさにより分別する方法、密度差により分別する方法等、さまざま
な方法が知られている。例えば、異なる密度を有する粒子群を密度差により分別する方法として
は、密度を調節した分離液を用いる沈殿平衡法や沈殿速度法、遠心分離法等の方法が挙げられる。
また、粒子として細胞を分別する場合には、細胞等の生体試料が有する固有の免疫学的な特性で
ある特異的な抗原抗体反応を活用し、磁気粒子や蛍光分子を結合させ、磁気細胞分別または蛍光
活性化細胞分別のような細胞分別機により分別する方法や、マイクロ流路を用い、電気泳動や凝
集剤等を用いた方法も検討されている(特許文献7を参照)。また、マイクロ流路で超音波やピ
ンチド流路を用いて密度に従って粒子群を分別する装置(非特許文献1,2を参照)および微粒
子群を分級する方法が開発されている。
ストークスの式に従い、一定方向に流れる分別液中を、より高い密度を有する粒子群は、分別流
路中の比較的下方向の領域を流れ、また、より低い密度を有する粒子群は、分別流路中の比較的
上方向の領域を流れる。つまり、密度の差に従って粒子群が重力方向に異なる挙動を示すため、
粒子径(サイズ)が同じであっても、異なる密度を有する粒子群を分別し回収することができる。
従って、粒子に重力負荷等の大きなストレスをかけずに、短時間で粒子群を容易に分別すること
ができ、分別した粒子群を容易に回収することができるという効果を奏する。この技術は卵子や
体外受精卵など、細胞の分別に応用できる。
 JP 2013-252510 A 2013.12.19
JP 2013-252510 A 2013.12.19
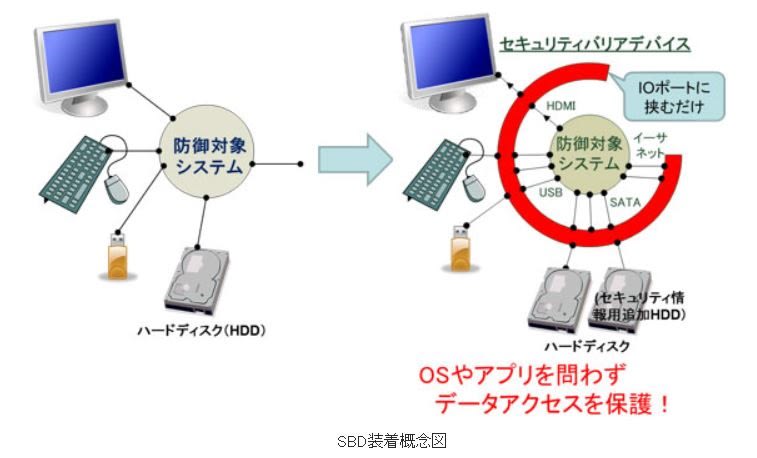
● 装着するだけで重要データを防御するセキュリティバリアデバイス
従来、パーティション単位での保護やファイルを保護領域に移動させるものはあったが、システ
ムの記憶装置に全く手を加えずにファイルやデータブロックを自由に指定して防御できるような
技術はなかったが、防御対象のシステムと周辺機器を繋ぐIOポートを中継する形で装着すること
で、セキュリティの強化を行う。
 JP 2008-271538 A 2008.11.6
JP 2008-271538 A 2008.11.6
【CO2分離回収の大幅な省エネ化技術】
セリウムの酸化物を用いた大きな表面積を持つ多孔質吸着体を安価で簡単に合成法で、これまで
の多孔質吸着体に比べて、2~4倍のCO2吸着量を実証、さらなる吸着性能向上も見込まれ、化学
吸収法に比べて約40%の省エネ効果が期待できるものであるという。
 CO2分離回収省エネ技術
CO2分離回収省エネ技術
【生命細胞の改変のいま】

ところで、生物とは何か? と問われたらどのように応えるのか。一番良さそうなのは「自己複
製を繰り返し増殖する生命体」という解答が妥当ではないだろうか。そう考えてみると、「再生
医療」とは、肥大化した脳を使い胎児期にしか形成されない人体の組織が欠損した場合にその機
能を回復させ自己増殖する人間の行動ということになるだろうか。その話はさておいて、その中
核的な「生命細胞学」の生物工学の昨今の進展について俯瞰してみよう。
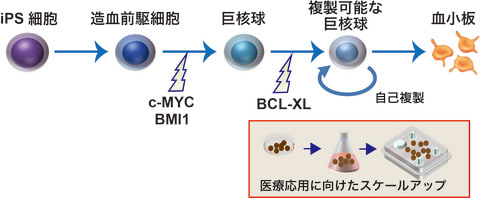
●ヒトiPS細胞から血小板を安定的に大量に供給する方法
従来の方法では、iPS細胞から輸血に必要な血小板量の100分の1程度しか作れなかったが、生体
外で自己複製し凍結保存が可能な不死化巨核球を誘導する方法を確立したことで、巨核球をスト
ックすることで血小板製剤の供給を安定化でき、今回の方法では、250億個の自己複製する巨核
球前駆細胞を使用して5日で血小板を得ることができ、培養する装置も、実験室レベルのシャー
レ(10mL)からバッグ(1~500L)にすることで複雑な設備を使わずに大量に培養することが出来
る。このシステムにより、日本人に多いHLA型のiPS細胞から血小板製剤を生産するための巨核球
のストックや、ドナーが見つかりにくいHLA型やその他の特殊な血小板型(HPA型)の患者への血
小板製剤の安定供給が可能となる。
※ Expandable Megakaryocyte Cell Lines Enable Clinically Applicable Generation of Platelets from Human
Induced Pluripotent Stem Cells Cell Stem Cell, 13 February 2014
 JP 2013-31428 A 2013.2.14
JP 2013-31428 A 2013.2.14
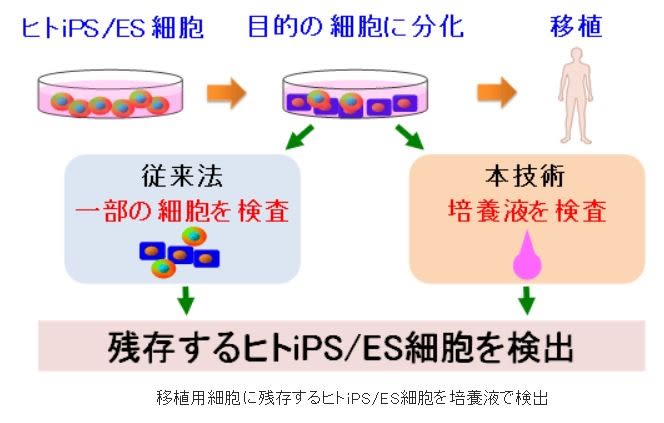
【再生医療に用いる細胞の安全性を培養液で検査】
産総研は、ヒトiPS/ES細胞から分化させて作製した移植用細胞は、未分化のヒトiPS/ES細胞が
残存する場合があり、この未分化ヒトiPS/ES細胞が腫瘍化する危険性があり、ヒトiPS/ES細胞
を医療に応用する際の最大の障壁となってたが、貴重な移植用細胞の一部を無駄にすることな
く調べることで、ヒトiPS/ES細胞の安全性を事前に把握することができる方法を開発したと公
表。今回開発した検出システムを用いると、多数の検体を迅速(3時間以下)に検査できる。
また、10 mLの培養液で1000万個の細胞を培養している場合、5000個(0.05%)以上のヒトiPS/
ES細胞が検出できる。移植用細胞中のヒトiPS/ES細胞の混入率を測定できるため、ヒトiPS/ES
細胞を用いた再生医療の安全性評価法として期待できる。
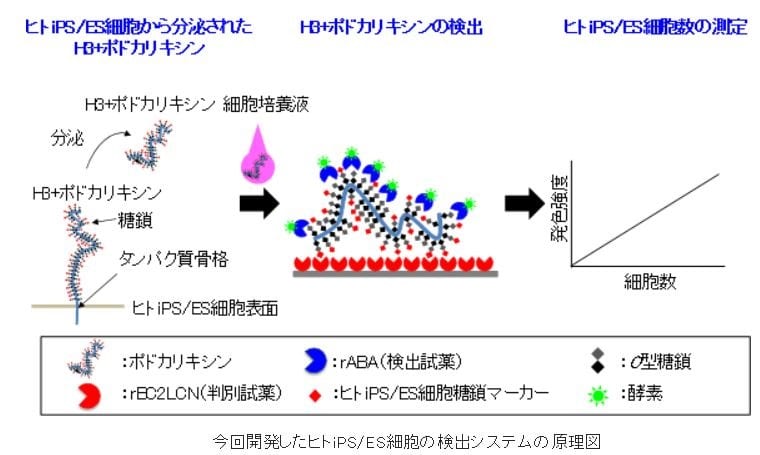
●要約
ヒトiPS/ES細胞に特徴的なO型糖鎖をもつポドカリキシン(H3+ポドカリキシン)が、さまざま
な種類のヒトiPS/ES細胞から培養液中に分泌されていることを見いだした。一方、ポドカリキ
シンは腎臓など他の組織にも存在するが、ヒトiPS/ES細胞に特徴的なH3+ポドカリキシンは調
べた限り通常の体細胞からは分泌されていない。培養液中のH3+ポドカリキシンを調べること
で、細胞自体を使わずにヒトiPS/ES細胞を検出できる。通常、タンパク質は特有のアミノ酸配
列を認識する抗体を用いて検出することが多いが、ポドカリキシンは多量のO型糖鎖で覆われ
た巨大なムチン様タンパク質であるために、抗体を用いることはできなかった。そこで、H3+
ポドカリキシンに多く存在する特徴的な糖鎖構造に着目し、そのO型糖鎖を認識するレクチン
を2種類用いて検出する新しいサンドイッチアッセイ系による検出システムを考案。詳細な方
法は以下の通り。
①H3+ポドカリキシンを認識するrBC2LCNを判別試薬として固定化した反応容器を準備する。
②1滴(50 µL)の細胞培養液を反応容器に入れ1時間反応させてH3+ポドカリキシンを吸着させ
る。
③洗浄して細胞培養液を除いた後、rBC2LCN とは別のO型糖鎖を認識する酵素標識rABAを、反応
容器に吸着したH3+ポドカリキシンと1時間反応させる。
④ 酵素標識rABAを発色させ、その発色強度を測定して、H3+ポドカリキシン量を決定する。
⑤ H3+ポドカリキシン量から、H3+ポドカリキシンを分泌したヒトiPS/ES細胞数を算出する。
※ JP 2013-39043 A 2013.2.28
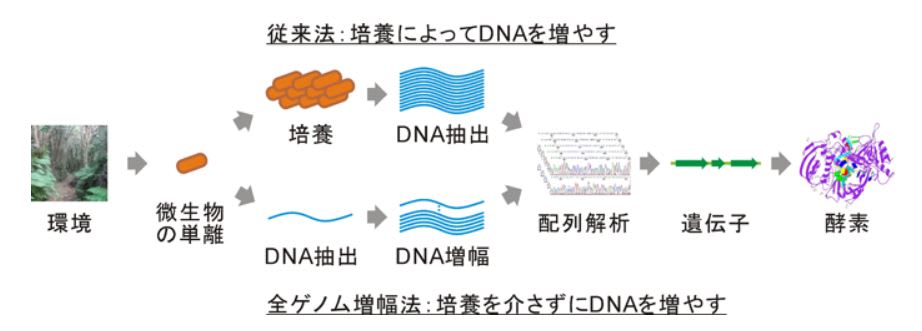
【微生物1個のDNAでも解析可能に】
農研機構は、微生物を培養して得られる遺伝子や培養産物から多くの有用物質が探索、発見さ
れてきたが、現代においても、培養できる微生物は全微生物の1%以下であり、99%以上の培
養できない微生物は有用物質の探索に活用されていない。そのため、微生物の遺伝子を、その
微生物を培養することなく、特殊なDNA合成酵素を用いて、培養できない微生物のDNAを大量複
製し、探索に必要な量のDNAを得ることが試みられているが、このDNA合成酵素には、試料DNA
が少ないほど、試料と無関係のDNAが複製されて、試料解析を困難にする問題があった。農研機
構は、高純度のDNA合成酵素を精製する方法を用いてこの問題を解決し、共同研究機関である関
東化学株式会社が本酵素を商業生産できる体制を構築することを公表。この酵素は、関東化学
株式会社から2014年度上半期に発売予定している。この発明により遺伝子の探索対象が大幅に
広がる。
●要約
全ゲノム増幅法で用いるDNA合成酵素は、組換え大腸菌で生産し精製する、生産に用いた大腸菌
のDNAが極わずかに混入し、この酵素で極小量の試料DNAを大量に増幅すると、混入した大腸菌
DNAが先に増幅(→これが試料と関係のないDNAが増える原因)。これを防ぐために、DNA合成酵
素から大腸菌DNAを除去する必要があるが、DNA合成酵素はDNAと強く結合する性質があり分離困
難だった。 そこで、大腸菌DNAを沈殿させて除去し、除去しきれないDNAはDNA合成酵素によっ
て複製されないように変性・分解する方法によって、大腸菌DNAを含まない高純度DNA合成酵素
を得る。この精製方法で調製したDNA合成酵素を全ゲノム増幅法に用いると、大腸菌の1個に含
まれるゲノムDNA量の約1/100に相当する微量な試料DNAも増幅できる。従来では困難であった
微量なDNAの全ゲノム増幅法が可能になり、培養できない微生物の遺伝子探索か可能となる。

※ Preparation of Phi29 DNA Polymerase Free of Amplifiable DNA Using Ethidium Monoazide, an
Ultraviolet-Free Light-Emitting Diode Lamp and Trehalose February 05, 2014
※ JP 2014-16344 A 2014.1.30
 JP 2014-16344 A 2014.1.30
JP 2014-16344 A 2014.1.30
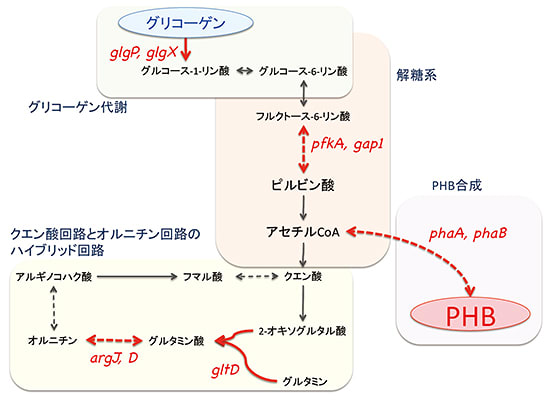
●ラン藻のバイオプラスチック生産が3倍増
理化学研究所は、Rre37タンパク質を細胞内で増やすとバイオプラスチック生産量が約2倍に増
加し、Rre37とSigEタンパク質を同時に増やすと生産量が約3倍に増加できることを解明したと
公表。このラン藻における代謝の制御機構を明らかにし基礎と応用の両面で研究を推進させたこ
とにより低コストでのラン藻培養法や回収法、効率的なPHBの抽出・精製法の開発が可能となる。
※ Pathway-Level Acceleration of Glycogen Catabolism by Response Regulator Rre37 in the Cyanobacte-
rium Synechocystis sp. PCC 6803". Plant Physiology , 2014,doi: 10.1104/pp.113.232025

【生物の起源 種とはなにか】
●一般的な感覚とは合致しない?
生物学的種概念は、有性生殖をするほとんどの生物においては、われわれのナイーブな
感覚とよく合致する。しかし問題がいくつかある。この種概念は基本的には、遺伝子交換
が可能な閉鎖群をもって種とする、と考えるため、無性生殖をする生物が扱えなくなるこ
とがひとつ。無性生殖生物については別の種概念を考える必要がある。もうひとつは、形
態や生活様式がはっきりと異なる生物集団であるのに、雑種が生じて遺伝子の交流が起き
るものがあることである。これは、ウマとロバの間にラバができるといった話とは遺う。
ラバには生殖能力はなく、ウマの集団とロバの集団で、遺伝子交換が起きることはなく、
ラバの存在は生物学的種概念に抵触しない。
テムプルトン(1989)によれば、アカガシワとクロガシワは同じ森の中で雑種をつ
くり、一部戻し交配が起こって遺伝子交換をするが、決して融合することはなく、はっき
り区別できる2つの集団に保たれているという。生物学的種概念を厳密に適用すると、ア
カガシワとクロガシワは同一種ということになるが、これはわれわれのナイーブな感覚に
反する。テムプルトンは、北米のオオカミとコヨーテの間でも同様なことが起こり、少な
くとも50万年以上、この状態で共存しているという。霊長類のヒヒ属ではさらに著しく、
菅原和孝(1990)によれば、マントヒヒとアヌビスヒヒ、アヌビスヒヒとキイロヒヒ、
キイロヒヒとチャクマヒヒの間で、それぞれ雑種ができるという。別属のゲラダヒヒとア
ヌビスヒヒの間にすら雑種らしき個体が観察されているという。馬渡峻輔(1994)は
マントヒヒとアヌビスヒヒは亜種レベルの種内変異と考えたほうがよいと述べているが、
この2つの集団は形態も生活様式も異なり、われわれのナイーブな感覚からすれば、明ら
かに別種である。
なぜ、こういうことになるかというと、生物学的種概念は、遺伝子の交換が起きるか起
きないかといった操作的あるいは事後的概念であって、存在論的意味合いが稀薄な概念だ
からである。そこで、遺伝子の交換を妨げている実際のメカニズムを実定し、これによっ
て種を定義しようと考えたのが、たとえば、配偶者認知システムによる種概念である(パ
タスン、1985)。単純に言えば、互いに同じ種であると認知し、交配を遂行できるメ
カニズムをもつ集団をもって種とする、という考えである。別の言い方をすれば、交尾前
隔離機構をもって種の定義とするということである。
隔離機構には交尾後隔離機構もあり、これは交尾が成立しても、この交尾を起点として
は遺伝子が未来に伝わらないなんらかのメカニズムのことである。この中で最も重要なの
は、雑種においては、父親由来の染色体と母親由来の染色体は、数や形が違うため、減数
分裂の際の対合(相同染色体がぴったりと合わさること)が起きず、結果的に機能的な配
偶子(卵や精子)がつくられないことである。もし、種になんらかの存在論的根拠がある
ならば、交尾後隔離機構の少なくとも一部は、細脳内における種の基本システムの違いに
より生じているとも考えられ、事は極めて重大であるが、これについては減数分裂も含め、
稿をあらためて論じたい。

●進化的運命共同体ではありえない
さて、話を元に戻す。さまざまな不都合があるとはいえ、生物学的種概念のような操作
的種概念が、配偶者認知システムといった実定的種概念よりも二股に受け入れられている
のは、種を進化の基本単位とみなしたいというところからくる。もし、今日の正統的な進
化論(ネオダーウィニズム)が主張するように、進化が遺伝子の突然変異と自然選択によ
って起こるならば(私自身はそういう考えに反対である)、実質的に遺伝子交換が可能な
閉鎖群は進化における運命共同体となるはずだ。なぜならばネオダーウィニストが主張す
る進化とは、通常、有性生殖プロセスを通じてある遺伝子が実質的な閉鎖群の中で多数に
なったり、少数になったり、絶滅したりすることにほかならないからだ。しかし、遺伝子
交換が可能な閉鎖群が必ずしもすべて同一の進化的運命共同体になるわけではない。
先はどのアカガシワとクロガシワについて考えてみる。もし、この2集団が遺伝子の交
換をすることによって融合するならば、この2集団は進化的には運命共同体だと考えてよ
い。この場合は生物学的種と進化的運命共同体は重なる。しかるにもし、この2集団がい
ずれ雑種をつくらなくなるか、雑種をつくり続けて遺伝子の交換をしながらも独自性を保
ち続けるのであれば、生物学的種と進化的運命共同体は異なるものとなる。
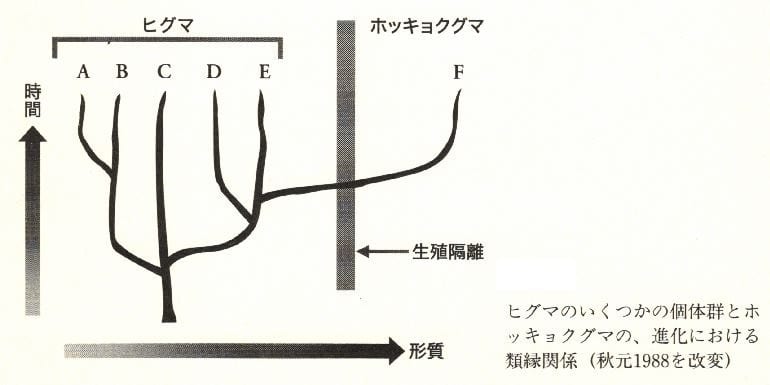
これについては、別のやっかいな問題がある。秋元信一(1988)によれば、ホッキ
ョクグマは、もっとも北方に進出したヒグマの個体群から2万年程前に分化して生じたと
いう。下図をみていただきたい。系統的には、ヒグマEはヒグマAよりもホッキョクグマ
Fに近縁である。A~Fは、さしあたってそれぞれ別の進化的運命共同体とみなしてよい。
これはもちろん生物学的種とは重ならない。系統関係を重視して、しかも生物学的種と系
統が異なるというジレンマを解決するやり方は2つある。ひとつは、ヒグマとホッキョク
グマを同一種にしてしまうやり方。もうひとつはA~Fの進化的運命共同体をそれぞれ別
種とみなすやり方である(クレイクラフト、1993)。
●生物自体があいまいなのだ
しかし、このどちらにしても、ここからはそもそも種は人間の認識論的概念であるとの
観点が抜け落ちてしまっている。もちろん、客観的に種が定義できれば、それでよいでは
ないかと考える人もいるだろう。たとえばA~Fの進化的運命共同体が未来にわたって他
と融合せず、分岐をするだけか、絶滅するだけであるならば、これをもって種とするとの
定義は一応つじつまは合う。しかし、進化的運命共同体が独自の進化をとげて、どこから
も文句の出ない独立種になるか、それとも他の集団と融合してしまうかは、そのときにな
ってみなければわからないのである。
さらに遺伝的隔離により区別される集団でさえ、隔離機構がなんらかの変異により消失
してしまい、近縁の種と融合してしまうことは十分考えられる。もちろん、種あるいはそ
れより上位の階級において、さしあたって不変のシステムが構想できれば、これをもって
生物の基本単位であると考えることはできるし、ここで詳しく述べる紙幅はないが、私が
主張する構造主義生物学は、基本的にはそのような構想を擁護する。
しかし、いずれにせよ、われわれがナイーブに認知する種は、厳密な存在論的根拠をも
っていないことは確かなようだ。テムプルトンの提唱する結合的種概念(Cohesion Conc-
ept)は厳密さや単純さを犠牲にして、われわれのナイーブな感覚に合わせようとの努力
であるといえる(下図)。テムプルトンは基本ニッチと遺伝的隔離の2つの指標に注目し、
その組み合わせで種を定義しようと考えた。
基本ニッチの異なるものは、遺伝的隔離のあるなしにかかわらず別種であり、基本ニッ
チが同一のものは、有性種で遺伝的隔離があるものだけが別種で、残りは同種である。こ
の定義に従えば、基本ニッチが同じ無性種は同種であり、アカガシワとクロガシワは別種、
ヒグマとホッキョクグマも別種であるが、系統の異なるヒグマどうしは同種になる。ずい
ぷんとあいまいな定義だが、ある意味ではやむをえない。定義のあいまいさは、生物のあ
いまいさの反映なのである。

池田清彦 著 『新しい生物学の教科書』 第1章
「種とはなにか」で池田清彦は「わたしたちは自然の中仁、形態や生活様式によって、他の生物
集団からははっきりと区別できる生物集団を認識することができる。これは種と呼ばれる。ニュ
ーギニアのアルファク山脈において、現地の狩人と生物学的訓練を受けたヨーロッパの科学者が
認識した鳥の種は、ほとんど全く同じだったとから考えて、種が認識できるのは、人間の基本的
な能力であることがわかる。もちろん種は人間め勝手な想像による架空の産物ではなく、多少と
も自然界に根拠をもつ存在である。多少ともと書いたのlは厳密には生物は系統をさかのぼれば
連続的な存在であるからだ。連続的なものが進化の結果、不連続なものとしてわれわれに認識可
能になったものが、現在見られる種であると考えられる。」「はっきりと不連続になった集団は
、形態、生理、発生、生態等々が違うことに加え、有性生物であれば、遺伝的に他の集団から隔
離iきれるので、遺伝的隔離という明確な種の定義を採用するレことができる。しかし、不連続に
なりつつある集団や、いったん不連続になりかけたものの再び融合しづつある集団に関ノしては、
厳密な定義を下すごとはできない。種に対して厳密な定義を下せないのは種が進化することの必
然の結果なのである。もしかしたら将来の科学者は、共通の生物学的システムといった何らかの
確固とした根拠に裏づけられた、生物のグルーピング法を見出すかもしれない。しかし、それが
現在われわれが認識している種と、一致するかどうか定かではない。」と意味深なことを指摘し
ている。
※「生物の起源 -細胞生命の起源-」、仲田崇志 2010.09.01

さても、「生物とはなにか」→「生物の起源とはなにか」との問いかけとはやっかいな問題を投
げかけ、わかった風にして泰然としているわたし(たち)に、無知を自覚させる。これは残件と
して暫く考えていくしかない。











 Wikipedia
Wikipedia
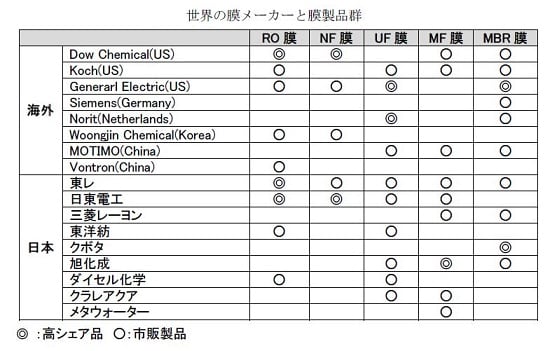
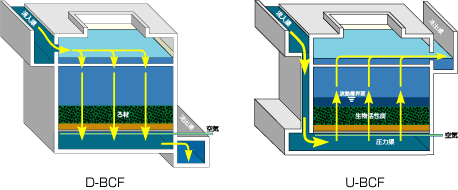
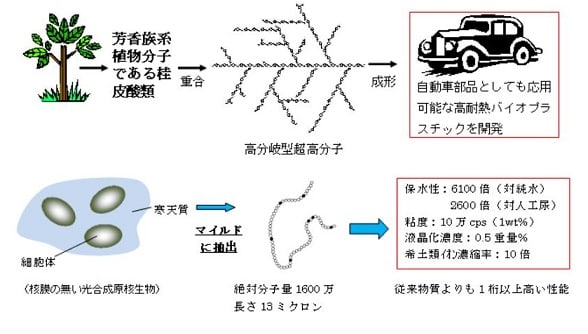
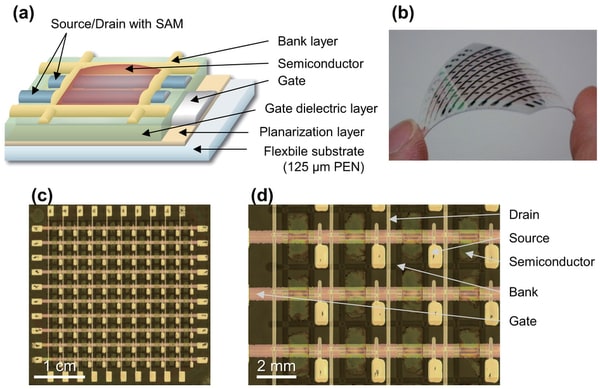
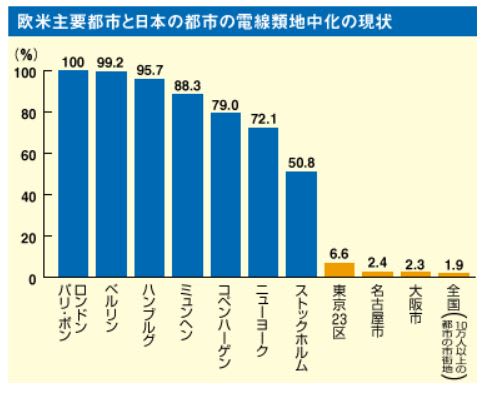
 特開2014-014739
特開2014-014739
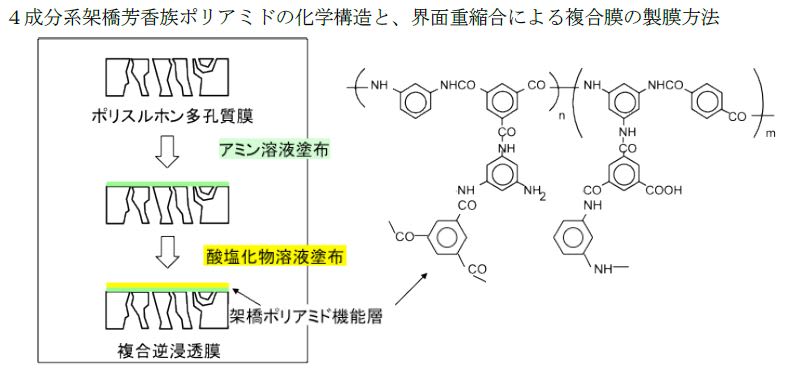
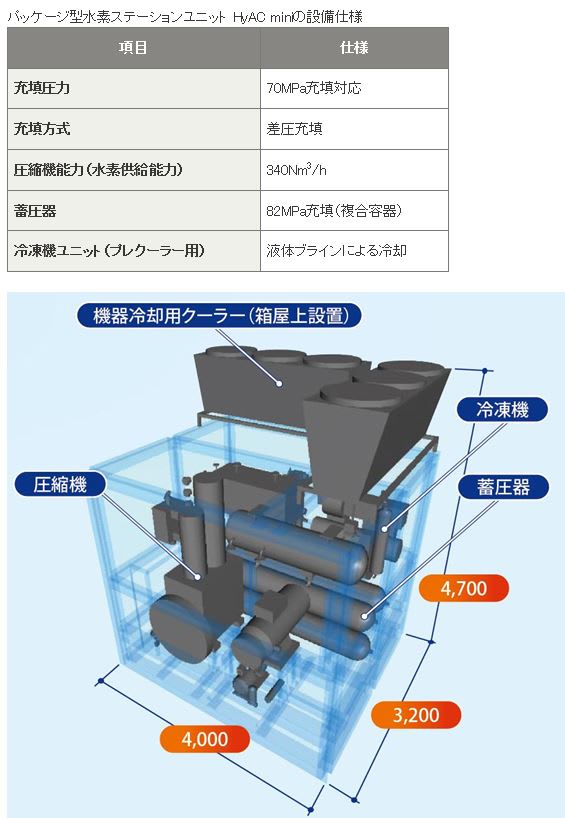
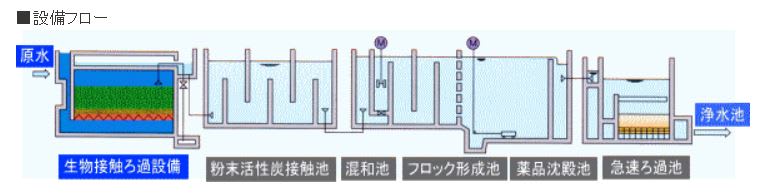
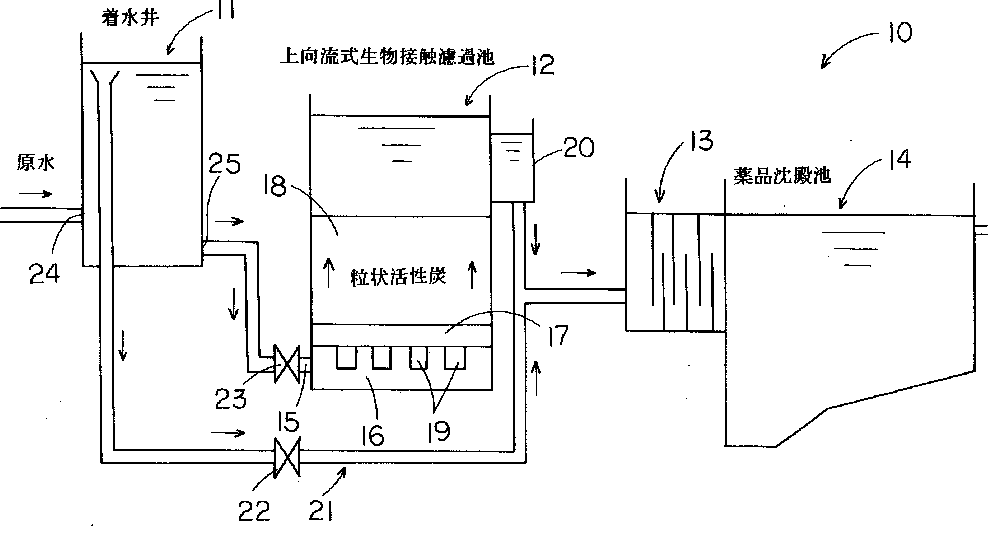
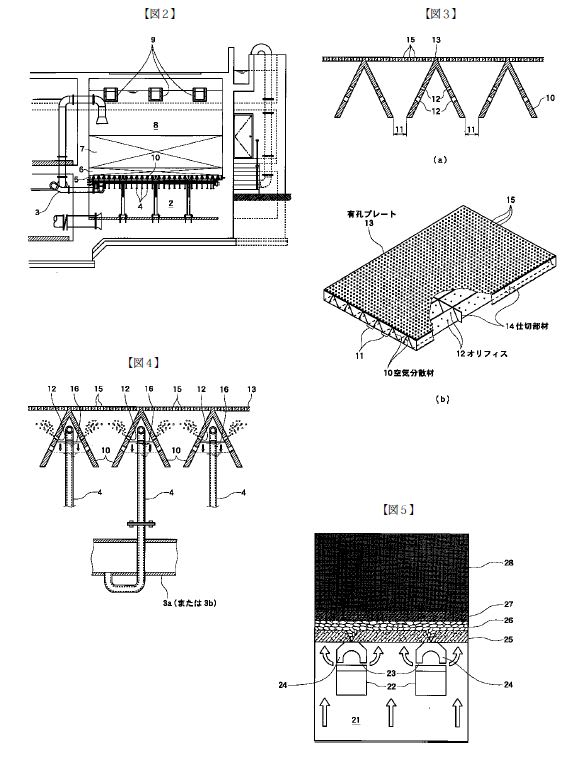
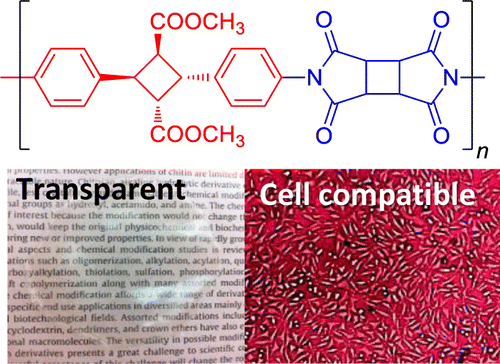
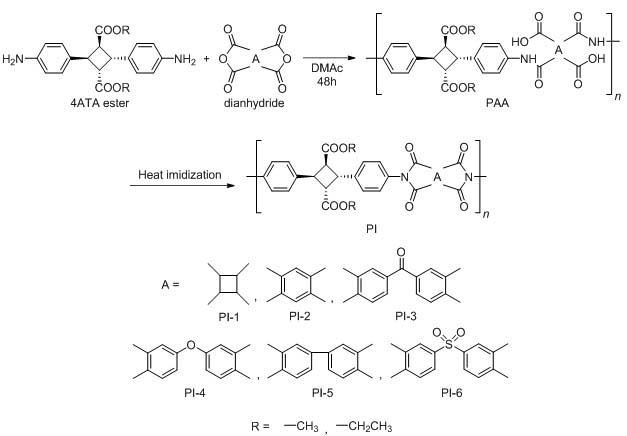
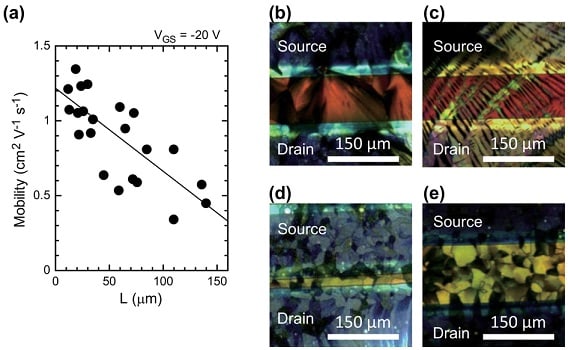
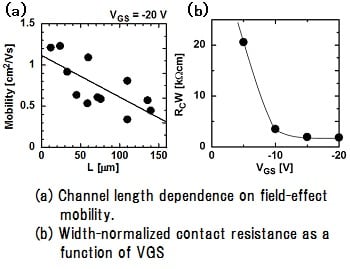
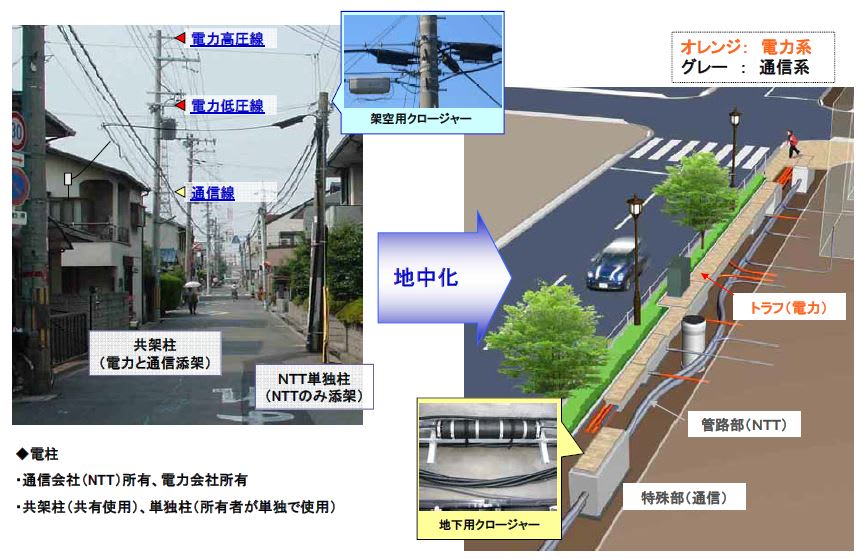
 特開2013-130218
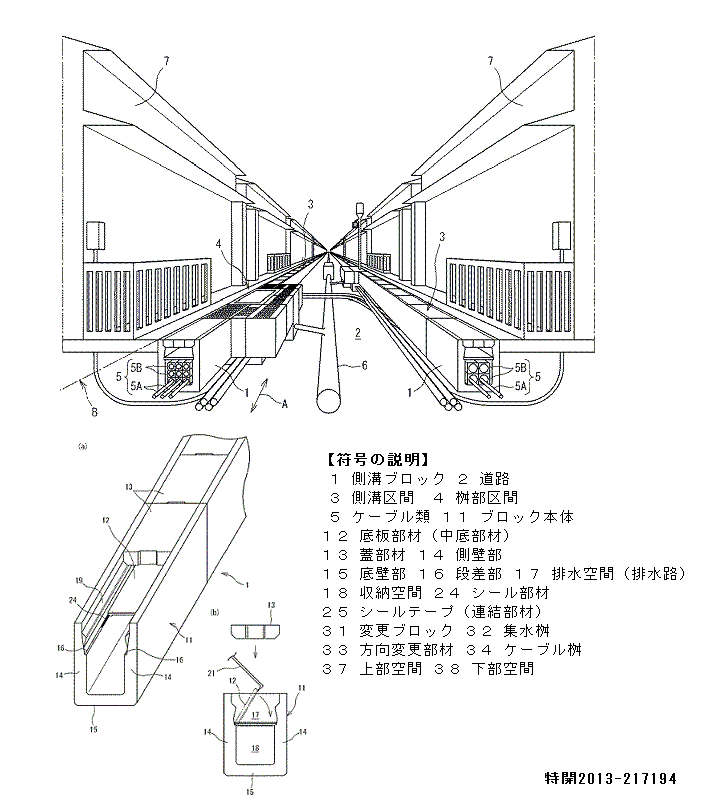
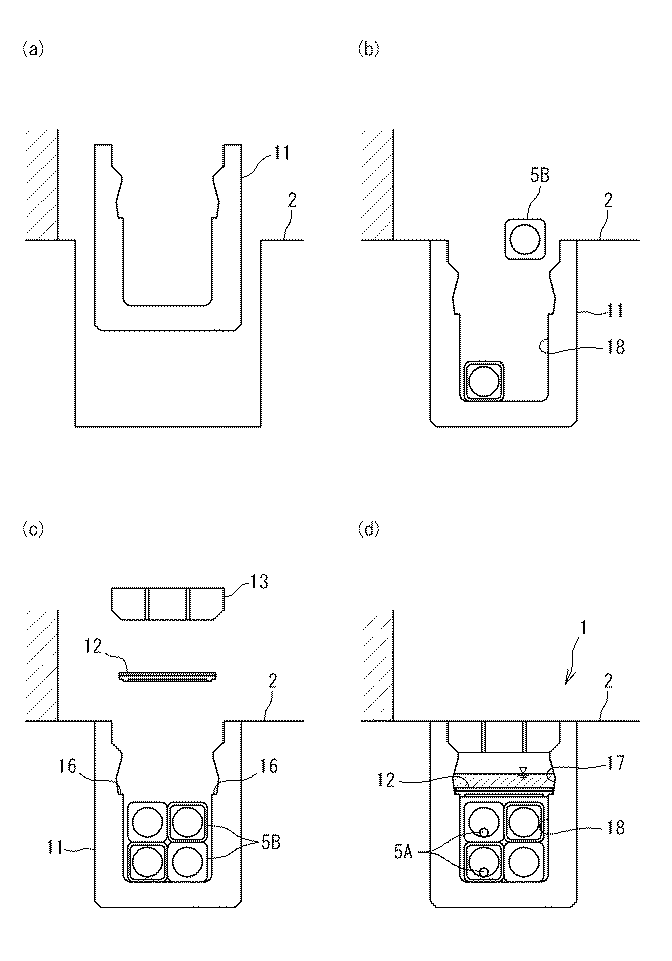
特開2013-130218 特開2013-224233
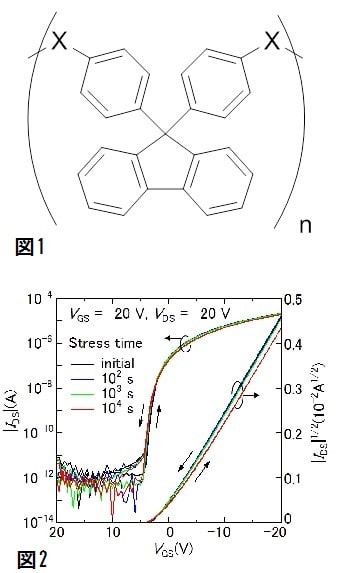
特開2013-224233



















































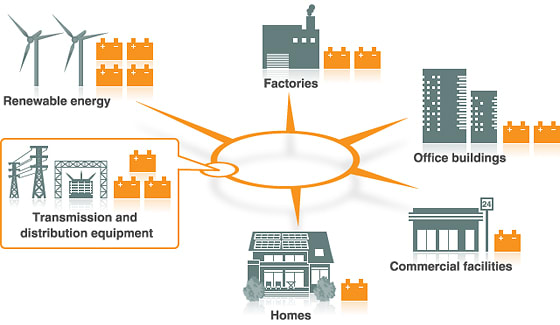



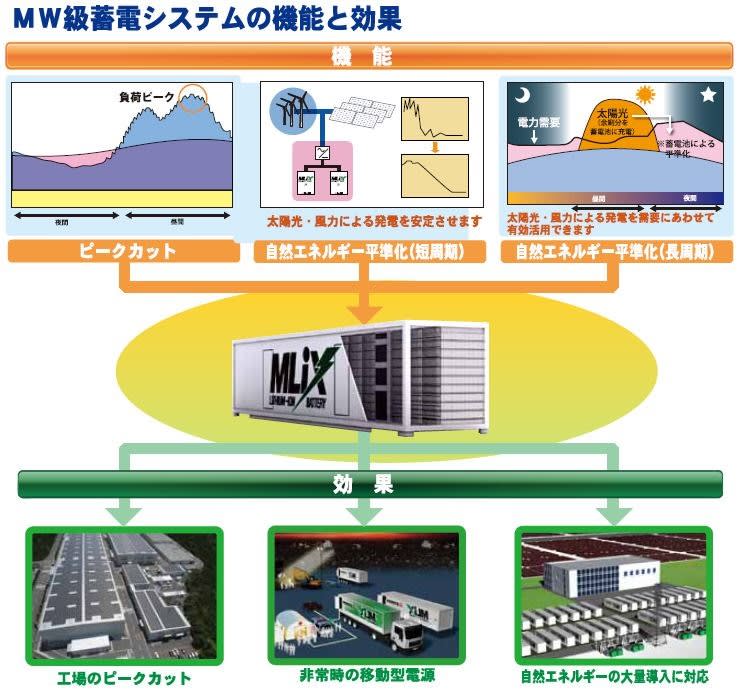
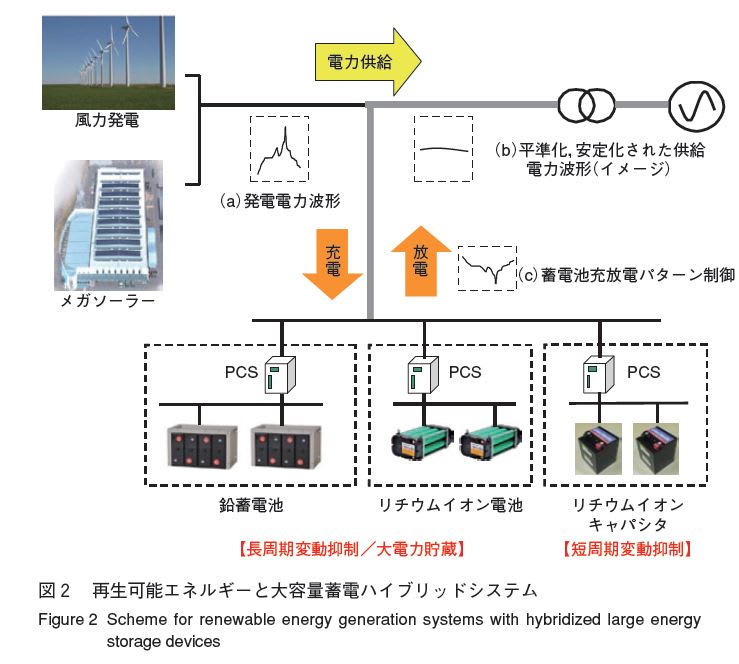



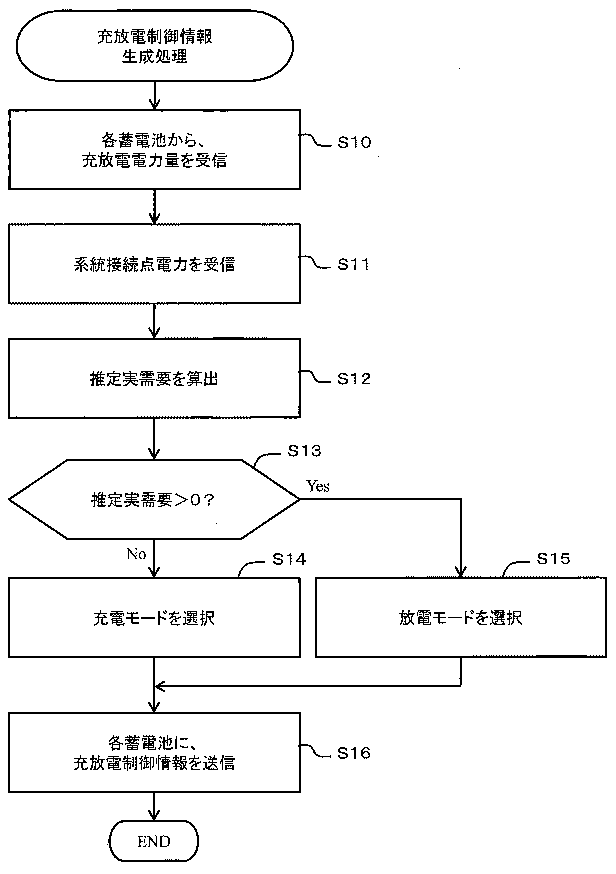





 今朝の一汁一菜
今朝の一汁一菜